Bioactive Materials Derived from Menstrual Blood Stem Cells Enhance the Quality of In Vitro Bovine Embryos
-
Amini , Mohammad Sobhan
-
Department of Animal Science, Science and Research Branch, Islamic Azad University, Tehran, Iran
-
Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
 Naderi, Mohammad Mehdi
Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran, Tel: +98 21 22432020; Fax: +98 21 22432021; E-mail: m.naderi@avicenna.ac.ir; mm.nadery@yahoo.com
Naderi, Mohammad Mehdi
Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran, Tel: +98 21 22432020; Fax: +98 21 22432021; E-mail: m.naderi@avicenna.ac.ir; mm.nadery@yahoo.com
-
Shirazi, Abolfazl
-
Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Aminafshar, Mehdi
-
Department of Animal Science, Science and Research Branch, Islamic Azad University, Tehran, Iran
-
Borjian Boroujeni, Sara
-
Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Pournourali , Mostafa
-
Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Malekpour , Ali
-
Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
Abstract: Backgrounds: The aim of this study was to determine whether the addition of bioactive materials derived from Menstrual Blood Stem Cells (MenSCs) to the oocyte maturation medium may improve the quality of bovine embryos in vitro.
Methods: MenSCs were collected from 6 healthy women (between 26 and 36 years old) and after 3 days of culture, their bioactive materials were frozen. The bovine Cumulus-Oocyte-Complexes (COCs) were aspirated from ovarian slaughterhouse and the oocytes with more than three layers of cumulus cells were cultured in vitro in media supplemented with (treatment) and without (control) 10% MenSCs’ bioactive materials. After IVM/IVF, the presumptive zygotes were cultured for 8 days.
Results: The blastocyst rate on day 8 in treatment group was higher than control (40.2±1.9 vs. 23±4.2.3, p=0.001). The ratio of Trophectoderm (TE) and Inner Cell Mass (ICM) (ICM/TE) cells was also greater in treatment group compared to control (30.3±2 vs. 14.9±1; p=0.001). The re-expansion of vitrified blastocysts, 24 hours after warming, in treatment group was higher than control (93.3±2.5 vs. 66.2±8.8; p=0.01). The expression of some genes related to pluripotency and implantation (OCT4, CDX2, and IFNT) were increased in treatment group compared to control (p<0/05).
Conclusion: In conclusion, the addition of MenSCs’ bioactive materials during in vitro maturation of bovine oocytes could improve the quantity and quality of bovine IVP embryos. Also, the expression of some genes associated with pluripotency and implantation in the blastocyst would be increased.
Introduction :
Since the production of the first calf derived from in vitro Fertilization (IVF) in 1981 1, significant advances have been made in improving the in vitro Production (IVP) of bovine embryos. However, the success rate in the terms of blastocyst rate varies between 30 and 40% 2-4. Furthermore, the quality of IVP embryos is lower than that of in vivo embryos in terms of morphology and cryo-survivability 5,6. Several factors are effective in development of pre-implantation stage embryos such as oocyte and sperm quality, protein and energy source, oxygen concentration, and the number of embryos per culture unit 2. Previous studies have shown that supplementing the embryo culture medium in vitro with different growth factors may reduce embryo apoptosis and degeneration and increase blastocyst quality (total cell number and hatching rate) in farm animals 7.
Stem cells have been known as a source of various growth factors that are able to regenerate themselves over long periods of time. The endometrial lining of the uterus has a remarkable capacity for regeneration. In 2007, the researchers from Medistem for the first time isolated a highly proliferating adherent cell population from menstrual blood with non-hematopoietic markers, termed as Menstrual Blood Stem Cells (MenSCs) 8. There are some multipotent markers in uterine stromal cells that are commonly found in bone marrow mesenchymal stem cells 9. These cells may have originated in the bone marrow, expressing C-kit cell surface markers (CD117) along with Oct-4 and SSEA-4 expression as important markers for determining multipotency, which have been shown in many pluripotent stem cells, including Embryonic Stem Cells (ESCs) 10-12.
Over the past decade, researchers have become more interested in using MenSCs because of their many advantages including: as an abundant and continuous source of growth factors, the possibility of preparing it through non-invasive procedure, high proliferative rate, low immunogenicity, and lack of ethical issues when compared with other source of stem cells 13,14. Moreover, MenSCs could stably multiply for at least 20 passages without mutations or visible abnormalities in vitro 8,15. This study was designed to evaluate the potential beneficial effects of bioactive materials derived from cultured MenSCs, during IVM, as a source of growth factors in promoting developmental indices of in vitro produced bovine embryos.
Materials and Methods :
All experimental procedures were reviewed and approved by Avicenna Research Institute, the Bioethics Committee (no 1910). All chemicals were purchased from Sigma chemical company (St. Louis, MO, USA), unless otherwise indicated in the text.
Oocyte collection and in vitro maturation (IVM): The bovine ovaries were collected from local slaughterhouse and transported to the laboratory in sterile saline supplemented with antibiotics (0.3 gr/L pen/strep) at 37°C within 2-3 hr. Extra tissues were removed and ovaries were washed with normal saline solution and all visible ovarian follicles with a diameter of 2-8 mm were aspirated using 18-gauge needle fitted with a 20-ml syringe. The Cumulus Oocyte Complexes (COCs) with more than three layers of cumulus cells and homogenous cytoplasm, were used for culture. About 10 COCs were cultured in 50-μl droplets of maturation media containing 475 μg/ml Bicarbonate Buffered M199, 105 μg/ml NaHCO3, 2 μg/ml sodium pyruvate (2´), 2.5 μg/ml Penicillin-Streptomycin, 0.05 IU/ml FSH, 0.01 IU/ml LH, and 10% Fetal Bovine Serum (FBS) in 65 mm culture dish overlaid with mineral oil for 24 hr at 38.5°C and 5% CO2 in air with 100% relative humidity. The COCs were randomly allocated and cultured into two groups: I) IVM medium supplemented with 10% FBS (control group) and II) IVM medium supplemented with 10% bioactive material derived from menstrual blood stem cell plus 5% FBS (treatment group).
In vitro fertilization (IVF): Frozen-thawed semen from a single bull was used for the in vitro fertilization of matured oocytes throughout the study. The matured COCs were subjected to in vitro fertilization in F-TALP media supplemented with 0.2 mM sodium pyruvate, 5 mg/ml Bovine Serum Albumin (BSA) (Fatty Acid free), 20 μg/ml heparin, and 5 μg/ml Penicillin-Streptomycin. Single frozen bovine semen straw (0.5 ml) was thawed at 37°C for 30 s and motile sperm was obtained by centrifugation based on pure sperm gradient (500 µl 40% over 500 µl 80%) at 500 g for 5 min. The supernatant was discarded and the sperm pellet was resuspended to a final concentration of 2×106 spermatozoa/ml using F-TALP medium. Progressively motile sperm from the supernatant solution of sperm pellet were placed as 50-μl droplets under mineral oil. The washed oocytes were then put into the 50-μl semen droplets (10 oocytes/50 μl droplets) and incubated for 18 hr at 39°C and 5% CO2 in air with 100% relative humidity.
In vitro culture of embryos (IVC): After 18 to 20 hr of IVF, to remove cumulus cells, the presumptive zygotes were vortexed for 2 to 3 min. The zygotes were then cultured in embryo culture media (SOF) supplemented with 2% (v⁄v) BME- essential amino acids, 1% (v⁄v) MEM-non essential amino acids, 1 mM glutamine and 8 mg⁄ml fatty acid free BSA at 39°C under conditions of 7% O2, 5% CO2 and 88% N2 in humidified air. On the third and fifth day of culture, 10% (v⁄v) charcoal stripped FBS was added to the medium. The osmolarity was maintained at 270 to 285 mOsmol. The percentage of cleaved embryos on day 3 and the percentage of total blastocysts on days 6, 7 and 8 were expressed on the basis of the number of oocytes at the onset of culture, and the percentage of expanded and hatched blastocysts on days 6, 7, and 8 was proportional to the total number of blastocysts on the same day.
Embryo cryopreservation: For embryo vitrification, fully expanded blastocysts (days 6 to 8) from each group with normal morphology were vitrified by Minimum Volume Cooling (MVC) method using a Cryotop device (Kitazato Supply Co, Toyko, Japan). Briefly, embryos were placed in basal medium (HEPES-TCM199+20%FBS) for at least 1 min and then placed in an equilibration solution comprised of 7.5% Ethylene Glycol (EG) and 7.5% Dimethyl Sulfoxide (DMSO) in basal medium for 12-15 min. After embryo shrinkage and return to the original size, embryos were exposed to vitrification solution (16.5% EG, 16.5% DMSO, 0.5 M sucrose in basal medium) for 30 s. The procedure was carried out at room temperature. Embryos were loaded on Cryotop and immediately immersed into liquid nitrogen. After 1 week, embryos were warmed directly in warming solution containing 1 M sucrose at 37.5°C for 1 min, followed by stepwise dilution of the cryoprotectants in solutions containing sucrose reducing concentrations (0.75 M for 3 min, 0.5 M for 3 min, and 0.25 M for 5 min, respectively). The warmed blastocysts were cultured for 24 hr in SOFaaBSA+10% CSS and the rate and speed of their re-expansion, survival, and hatching rate were assessed and compared among experimental groups. Embryos that were re-expanded within 8 hr after warming were considered as fast re-expanding embryos and those that were re-expanded between 8-16 hr after warming were considered as slow re-expanding ones. The survival rate of vitrified-warmed blastocysts was assessed using an inverted microscope at 400× magnification. The morphological integrity of the Trophoectoderm (TE; the ectoderm from which the trophoblast develops) and Inner Cell Mass (ICM) (equally shaped and sized), the capability of warmed blastocysts to re-expand, and the extent of dark and necrotic areas were assessed. The hatched blastocysts were used for counting embryonic and apoptotic cell numbers.
Preparation of MSCs and its bioactive materials: The bioactive materials of MSCs were prepared in Tissue Engineering Department at Avicenna Research Institute. About 3-5 ml of the menstrual blood from 6 healthy women (between 26 and 36 years old) were collected by the donors who signed the consent form, approved by the medical ethics committee of Avicenna Research Institute. The menstrual blood was collected using Diva Cup (Lunette, Stockholm, Sweden) on the second day of menstruation and immediately transferred to the laboratory. Then, collected menstrual blood cell was decanted into the tubes containing Phosphate Buffered Saline (PBS), supplemented with 0.5 mM ethylene diamine tetra acetic acid (EDTA), 2.5 mg/ml fungizone, 100 U/ml penicillin, and 100 mg/ml streptomycin. Mononuclear cells were removed using Ficoll-Hypaque separating system. Cell pellets were washed and suspended in complete DMEM-F12 and were cultured in polystyrene 75 cm2 tissue culture flasks, and incubated at 37°C in a humidified 5% CO2 incubator. Non adherent cells were washed away after 1-2 days of incubation. The culture medium was refreshed twice a week. Cells were passaged with Trypsin–EDTA after they became at least 70% confluent. The collected culture media of passage three was centrifuged in 1200 rpm for 5 min, then the supernatant (bioactive material) stored and kept at -80°C until use.
Embryo differential staining: For staining of Trophectoderm cells (TE), blastocysts were incubated in 0.2% Triton X-100 for 20 s and immediately transferred into 30 µg/ml Propidium Iodine (PI) solution for 1 min. For staining ICM, the blastocysts were incubated in ethanol containing 10 µg/ml bisbenzimide (Hoechst) for 15 min on ice and were then mounted on glass slides and examined under an epifluore-scence microscope 16.
Expression of Nanog, Bax, OCT4, CDX2, and IFNT genes: The primers were designed by oligo 7, and represented in table 1. In order to extract the RNA, 5 embryos (day 8) were washed at least 5 times in PBS, then transferred to 1.5 ml micro tube. The embryos were subjected to 3 times freeze-thaw process, from 37°C water to liquid nitrogen 17. The concentration of RNA was assessed by NanoDrop, and the extracted RNA was then stored at -80°C. First-strand cDNA was synthesized from the total amount of RNA using the cDNA synthesis kit (Takara, USA), following the manufacturer's instructions. The Reverse Transcriptase (RT) is a modified MMLV-derived reverse transcriptase and the Reaction Mix contains both oligo (dT) and random primers. After the RT reaction and RT control with primers for H2A gene as a house keeping gene, the cDNA was 2.5 times diluted in 10 mM Tris HCl at pH=8.0 and finally the data were analyzed by REST7.
Statistical analysis: Data were collected over at least four replicates. All data were expressed as mean±standard error of the mean (SEM). Differences among groups were analyzed by one-way ANOVA followed by post hoc Fisher LSD test using SigmaPlot (version 11.0). When equal variance test failed, the treatments were compared by Student-Newman-Keuls Method. When normality test failed, the Kruskal-Wallis ANOVA was applied. Differences were considered significant when p≤0.01.
Results :
Effect of MenSCs’ bioactive materials on bovine embryo development: There was no significant difference in cleavage and total blastocyst rates on day 7 between treatment and control groups. However, the difference, in total blastocyst rate on day 8 was significant between the treatment and control groups (40.2±1.9 vs. 23±4.2.3 respectively; p=0.001) (Table 2).
Effect of MenSCs’ bioactive material on blastocyst cell number: There was a significant difference in the number of ICM between treatment and control groups (31.7±2.5 vs. 18.2±3.9 respectively; p=0.005). The ICM to the TE and ICM to the total cell ratios between treatment and control groups were also different (Table 3).
Effect of MenSCs’ bioactive materials on embryo cryotolerance: There were no significant differences between bioactive materials and control groups in terms of hatching rates at 24 or 48 hr after warming the vitrified embryos (Table 4). However, the re-expansion rate 24 hr after warming was significantly increased in the bioactive material group compared to the control (93.3±2.5 vs. 66.2±8.8 respectively; p=0.01). Re-expansion after 48 hr of warming was also different between bioactive material and control groups (98.4±1 vs. 68.5±11.5 respectively; p=0.02).
Effect of MenSCs’ bioactive materials on expression of OCT4, CDX2, IFNT, Nanog, and Bax genes: The expression of OCT4, CDX2, and IFNT genes in the bioactive materials group was increased compared to the control. However, there was no significant difference between treatment and control groups for Nanog and Bax gene expression (Figure 1).
Discussion :
As known, higher oocytes developmental competence ensures a higher number of high quality blastocysts 18. It is also evident that the quality of embryos has been affected by the culture condition by which they were exposed 19,20. In the current study, the addition of bioactive materials derived from MenSCs in the oocyte maturation medium significantly increased the rate of blastocysts on day 8. Moreover, the number of ICM, ICM to TE, and ICM to total cells ratios, which are important criteria to assess embryo quality, were greater in treatment group compared to the control 21. The low ICM/TE ratio and cells number and aberrant allocation of ICM and TE cells in embryos at preimplantation stages may cause placental abnormalities and early fetal loss 22. In our study, the rate of expanded blastocysts in treatment group (bioactive materials) was significantly higher than control on days 7 and 8 of culture. Conversely, the hatching rate on day 8 was slightly higher in the control group compared to the bioactive material group (27.3 vs. 18.1%). Previous studies have indicated that MenSCs express some pluripotency markers including Oct-4, SSEA-4, Nanog, and c-kit and secrete a variety of angiogenic, anti-apoptotic, and supportive factors such as Vascular Endothelial Growth Factor (VEGF), Insulin-like Growth Factor 1 (IGF-1), Transforming Growth Factor β (TGF-β), Stem Cell Factor (SCF), Granulocyte Macrophage Colony-Stimulating Factor (GM-CSF) 23, and various growth factors such as Epidermal Growth Factor (EGF) 24, Nerve Growth Factor (NGF) 25, Platelet Derived Growth Factor (PDGF) 26, and Fibroblast Growth Factor (FGF) 27 that can affect the quality of in vitro produced embryos. As a result, the beneficial effect of bioactive materials derived from MenSCs can be related to several growth factors produced by MenSCs.
There are also several studies indicating that Leukemia Inhibitory Factor (LIF), one of the growth factors secreted by MenSCs, increased blastocyst and hatching rates and the total blastocyst cell numbers in buffalo 28. Regarding VEGF secreted by MenSCs, its supplementation on days 4 to 7 of porcine embryo culture has been shown to improve blastocyst formation and its total cell number 29. In bovine, in addition to the positive effects of VEGF on nuclear and cytoplasmic maturation of oocyte 30, this growth factor is involved in cell proliferation and migration, implantation of embryos, increasing cleavage and blastocyst rates, and the metabolic activity of the trophoblasts in an autocrine fashion 31. It has been shown that in cattle the combination of FGF, GM-CSF, and LIF secreted by MenSCs significantly increases embryonic development from the expanded blastocyst to the hatched blastocyst stage 32.
Regarding embryo cryopreservation, in our study, the rate of re-expansion of vitrified blastocysts was significantly increased 24 and 48 hr after warming in the treatment group compared to the control. In fact, the secreted bioactive materials increased the re-expression rate by 27 and 30% after 24 and 48 hr, respectively. As known, the degree of re-expansion is a significant predictor of live birth, comparable to the ICM/TE ratio in fresh and vitrified/warmed blastocyst 21.
In our study, a significant increase in the expression of OCT4, CDX2, and IFNT genes was observed in the treatment group compared to the control. In the study of Nichols et al although the addition of MenSCs’ bioactive materials in oocyte maturation media had no effect on the expression of Nanog and Bax genes in blastocysts, but there was a significant increase in the expression of OCT4, CDX2, and IFNT genes in the treatment group compared to the control group. It has been shown that Nanog, OCT4, and SOX2 genes have potential role in regulating gene expression and are essential for maintaining the pluripotency of embryonic stem cells and ICM functions. OCT4 as a transcription factor gene is a member of POU family that is found in the promoter and enhancer regions of many genes. OCT4 also regulates the expression of Nanog, SOX2 and other genes, and can modulate cell fate in the early stages of embryo development. In our study, the expression of Cdx2 that is responsible for embryo compaction and TE lineage formation was increased in treatment group compared to control. On the other hands, despite the lack of difference in Bax expression, the expression of other genes increased after the oocytes were exposed to the bioactive materials and showed that the bioactive material has a significant effect on blastocyst development 33.
Conclusion :
In conclusion, by adding MenSCs’ bioactive material to the IVM medium, the developmental competence of cattle oocyte can be improved. This is manifested by an increase in the quantity and quality of the blastocyst as well as an increase in the expression of some genes associated with pluripotency and implantation in the blastocyst.
Acknowledgement :
Thanks to Mrs. Maryam Darzi (Tissue Engineering Department at Avicenna Research Institute), Mrs. Haleh Edalatkhah (Reproductive Genetic Department, Avicenna Research Institute), Mrs. Maryam Salimi (Reproductive Biotechnology Research Center, Avicenna Research Institute), Mr. Ali Sarvari (Reproductive Biotechnology Research Center, Avicenna Research Institute) and Mrs. Bahareh Behzadi (Reproductive Biotechnology Research Center, Avicenna Research Institute) for providing us technical supports.
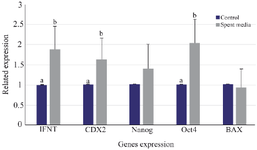
Figure 1. Effects of MenSCs’ bioactive materials on expression of Nanog, Bax, OCT4, CDX2, and IFNT genes.
|
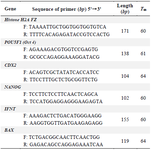
Table 1. The primers sequences used in Real Time PCR
|
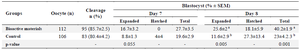
Table 2. Effect of supplementing embryo culture medium with MenSCs’ bioactive materials on bovine embryo development
a, b) The numbers with different uppercase letters at the same column are significantly different (p<0.05).
|
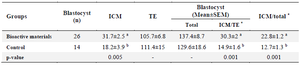
Table 3. Effects of MenSCs’ bioactive materials on blastocyst cell number
* Data are presented as (% ± SEM).
a, b) The numbers with different uppercase letters at the same column are significantly different (p<0.05).
ICM: Inner Cell Mass, TE: Trophectoderm.
|
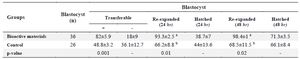
Table 4. Effect of MenSCs’ bioactive materials on re-expansion and hatching rates of vitrified/warmed embryos after 24 and 48 hr
a, b) The numbers with different uppercase letters at the same column are significantly different (p<0.05).
Data are presented as (% ± SEM).
|
|