Luteolin-Treated 4T1 Cell-Derived Exosomes as Novel Antiproliferative Agents In Vitro and In Vivo
-
Heidari, Pardis
-
Faculty of Veterinary Sciences, SR.C., Islamic Azad University, Tehran, Iran
-
 Hosseini, Razieh
Department of Veterinary Basic Sciences, SR.C., Islamic Azad University, Tehran, Iran, Tel: +98 9173059160; Fax: +98 21 44865166; E-mail: razieh.hosseini@iau.ac.ir, Hosseini_945@yahoo.com
Hosseini, Razieh
Department of Veterinary Basic Sciences, SR.C., Islamic Azad University, Tehran, Iran, Tel: +98 9173059160; Fax: +98 21 44865166; E-mail: razieh.hosseini@iau.ac.ir, Hosseini_945@yahoo.com
-
Nasihatgar , Nika
-
Faculty of Veterinary Sciences, SR.C., Islamic Azad University, Tehran, Iran
Abstract: Background: Breast cancer is the most widespread malignancy among women worldwide. Luteolin, a flavonoid, has demonstrated anti-cancer effects by triggering apoptosis in tumor cells. Exosomes are gaining much attention for cancer therapeutic approaches due to multitude of beneficial effects. This study is aimed to investigate the possible potential of exosomes derived from luteolin-treated 4T1 cells to ameliorate tumor in comparison to luteolin treatment only.
Methods: In this study, 4T1 cell culture was exposed to luteolin. Following exosome extraction, they were characterized using field emission scanning electron microscopy, dynamic light scattering and western blot analysis. MTT assay was performed in order to evaluate cell viability after exposure to different concentrations of luteolin and exosomes. An in vivo breast cancer model was induced via subcutaneous injection of 4T1 cells to the BALB/C mice. After 14 days, tumor volume was measured, and expression of RhoA and ERK mRNAs were quantified by Real Time PCR.
Results: The MTT assay demonstrated that exosomes from luteolin-treated 4T1 cells at a concentration of 320 μg/μl reduced cell viability by approximately 70% in a dose-dependent manner. Tumor volume in the exosome-treated group decreased by 57% relative to the tumor group, while the luteolin-treated group demonstrated a 39% reduction. Furthermore, RhoA gene expression was substantially downregulated in the exosome-treated group, and exosomes were more effective than luteolin in reducing ERK gene expression.
Conclusion: Exosomes derived from luteolin-treated 4T1 cells effectively suppress breast cancer cell growth by reducing 4T1 cell viability and by decreasing tumor volume and downregulating tumor-associated genes RhoA and ERK. These results propose a novel therapeutic strategy for breast cancer, highlighting the promising potential of exosomes as an efficient drug delivery system.
Introduction :
Breast cancer is the leading cause of tumor-related morbidity and mortality among women globally 1. Epidemiological evidence suggests that approximately 90–95% of cancer cases are associated with lifestyle and environmental factors, while only 5–10% are attributable to inherited genetic abnormalities 2. Chemotherapy and radiotherapy remain the mainstay treatment modalities for breast cancer; however, they are frequently associated with significant adverse effects that negatively impact patients’ quality of life 3. Despite substantial advances in technology and pharmaceutical interventions over the past two decades, cancer remains a major global health challenge 4. Consequently, there is a growing effort to develop alternative therapeutic strategies for patients who experience both physical and psychological complications as a result of conventional treatments.
Natural products and isolated bioactive compounds derived from medicinal plants have been shown to possess both cancer-preventive and therapeutic properties 3. Among these, flavonoids—a diverse class of polyphenolic compounds—have attracted considerable interest in cancer research, particularly in the context of breast malignancies. Flavonoids have been reported to induce apoptosis, inhibit cell viability and proliferation, and modulate critical signaling pathways involved in breast cancer progression 5,6. Luteolin (3′, 4′, 5, 7-tetrahydroxyflavone), a naturally occurring flavonoid abundant in various fruits and vegetables, is widely recognized for its anticancer properties 7. Its thera-peutic effects are primarily attributed to its ability to induce apoptosis in malignant cells and to inhibit tumor cell proliferation in both in vitro and in vivo models 8. Furthermore, luteolin has been shown to induce cell cycle arrest at different stages, thereby effectively limi-ting cancer cell proliferation and metastatic potential 9,10.
Exosomes are small extracellular vesicles, typically 30–150 nm in diameter, that play a crucial role in intercellular communication. They transport a wide range of bioactive molecules, including proteins, nucleic acids, and lipids 11. Moreover, exosomes exhibit several advantageous properties, such as high biocompatibility, low immunogenicity, structural stability, favorable pharmacokinetics, efficient biodistribution, and effec-tive cellular uptake, which make them promising can-didates for anticancer therapies 12. Indeed, exosomes can be engineered to deliver specific cargos that target oncogenic signaling pathways, thereby enhancing their therapeutic efficacy in cancer treatment 13,14. Relying on the aforementioned facts, this study explores the combined impact of luteolin and exosome on prevention of breast cancer progression. In this regard, anti-cancer effect of exosomes derived from luteolin-treated 4T1 cells was evaluated both in vitro using MTT assay and in vivo using tumor volume measurement and expression of RhoA and ERK genes.
Materials and Methods :
Cell line and cell culture: The 4T1 mouse breast cancer cell line was obtained from the Pasteur Institute (Tehran, Iran). Cells were maintained in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% Fetal Bovine Serum (FBS) and antibiotics (penicillin 100 U/ml and streptomycin 100 μg/ml) at 37°C in a humidified atmosphere containing 5% CO₂. The culture medium was refreshed every two days, and cells were passaged upon reaching approximately 85% confluency. Following the third passage, cells were seeded into appropriate culture plates and enumerated for subsequent experiments.
Exosome extraction: Exosomes were isolated from luteolin-treated 4T1 cells. Cells were cultured in medium supplemented with 10% exosome-depleted FBS, obtained by overnight ultracentrifugation. After 48 hr of incubation, the conditioned medium was collected, and exosomes were isolated using sequential centrifugation steps at 300×g for 10 min, 2,000×g for 10 min, 10,000×g for 30 min, and 110,000×g for 2 hr. The resulting exosome pellet was resuspended in Phosphate-Buffered Saline (PBS), and the protein concentration was determined using the Lowry assay (Bio-Rad, USA).
Field Emission Scanning Electron Microscopy (FE-SEM) and Dynamic Light Scattering (DLS): Exosome morphology was characterized using Field Emission Scanning Electron Microscopy (FESEM) and Dynamic Light Scattering (DLS). Exosome size was determined by DLS using a laser wavelength of 532 nm. For FESEM analysis, exosome samples were resuspended in 1.3% paraformaldehyde, and a drop of the suspension was placed onto a clean coverslip. The samples were then washed three times with deionized water and air-dried overnight at room temperature. Subsequently, all samples were coated with a 2-5 nm layer of gold by sputtering and examined using a Mira 3-XMU scanning electron microscope (TESCAN, Czechia) operated at an accelerating voltage of 20 kV.
Western blot assessment: Exosomal proteins were extracted using RIPA lysis buffer (Cytomatin Gene, Iran) supplemented with 1% protease inhibitor cocktail (Kia Zist, Iran). Equal amounts of protein were separated on a 10% SDS-PAGE gel (Sigma-Aldrich, Germany) and subsequently transferred onto a Polyvinylidene Difluoride (PVDF) membrane (Sigma-Aldrich, USA). The membrane was incubated with primary antibodies against CD63 and β-actin (Santa Cruz Biotechnology, USA), followed by horseradish peroxidase–conjugated secondary antibodies (Bio-Rad, USA). Protein bands were detected using an enhanced chemiluminescence reagent (Parstous, Iran), and band intensities were quantified using ImageJ software.
Cell viability analysis: Cell viability was assessed using the MTT assay. Briefly, 1×10⁴ cells were seeded into 96-well plates and allowed to adhere for 24 hr. The cells were then treated with increasing concentrations of luteolin (5, 10, 20, 40, and 80 μM) or exosomes (20, 40, 80, 160, and 320 μg/ml) for 48 and 72 hr. Following treatment, 100 μl of 0.1% MTT solution was added to each well, and the plates were incubated for an additional 4 hr. The supernatant was subsequently removed, and 100 μl of Dimethyl Sulfoxide (DMSO) was added to solubilize the formed formazan crystals. Absorbance was measured at 490 nm using a spectrophotometer. All experiments were conducted in triplicate.
Animal model: All animal experiments were conducted in accordance with the ethical guidelines approved by the Ethics Committee of Islamic Azad University, Science and Research Branch. A total of 24 female BALB/c mice, weighing approximately 20±2 g, were obtained from the Laboratory Animal Center of the Pasteur Institute. The animals were housed individually in cages with separate food and water containers and were provided ad libitum access to standard laboratory chow and water. Mice were maintained under controlled environmental conditions, including a 12-hr light/dark cycle, 50% relative humidity, and an ambient temperature of 23°C, for one week prior to experimentation to allow acclimatization and reduce stress.
A murine breast cancer model was established by subcutaneous injection of 5×10⁵ 4T1 cells suspended in 50 μl of phosphate-buffered saline into the right flank of each mouse. Treatment was initiated once tumors became palpable. The animals were randomly allocated into four groups: a control group, an untreated tumor group, a luteolin-treated group receiving intraperitoneal injections of luteolin (50 mg/kg; Sigma-Aldrich, USA) four times over a two-week period, and an exosome-treated group administered exosomes (20 μg in 100 μl) intraperitoneally on the same schedule. Tumor volume was measured every three days using calipers on days 1 and 14. At the end of the two-week treatment period, mice were euthanized, and tumors were excised for further analyses.
Gene expression analysis by real-time PCR: Total RNA was isolated from tumor samples stored at -80°C using TRIzol reagent (Kiazist, Iran). Complementary DNA (cDNA) was synthesized by reverse transcription in accordance with the manufacturer’s instructions using a cDNA Synthesis Kit (Parstous, Iran). Quantitative real-time PCR (qRT-PCR) was performed to evaluate the expression levels of the RhoA and ERK genes using SYBR Green PCR Master Mix (Addbio, South Korea). Primer sequences were designed using the NCBI Primer-BLAST tool and are presented in table 1. GAPDH served as the internal reference gene. Relative gene expression levels were normalized to GAPDH and calculated using the comparative 2^−ΔΔCt method. All qRT-PCR reactions were carried out in triplicate.
Statistical analysis: Data obtained from three independent experiments are expressed as mean±Standard Deviation (SD). Differences among groups were analyzed using analysis of variance (ANOVA) with GraphPad Prism software version 9.1.1. A p value of less than 0.05 was considered statistically significant.
Results :
Characterization of exosomes: DLS analysis indicated that the average diameter of 4T1-derived exosomes was approximately 120 nm (Figure 1A). As shown in figure 1B, the isolated vesicles exhibited a spherical morphology consistent with the characteristic features of exosomes. In addition, the particle population was relatively monodispersed, with sizes predominantly below 100 nm.
Expression of CD63 marker in 4T1-derived exosomes: Western blot analysis was performed to confirm the presence of exosomes in the isolated supernatant and to assess CD63 expression. The analysis demonstrated positive staining for CD63, a well-established exosomal marker. β-Actin was employed as a loading control to normalize protein levels, ensuring uniform sample loading across the gel (Figure 2).
Exosomes substantially reduce the survival of 4T1 cancer cells relative to luteolin MTT assay results indicated that luteolin at 80 μM and exosomes derived from luteolin-treated 4T1 cells at 320 μg/μl reduced cancer cell viability by 60 and 70%, respectively. Notably, the exosomes induced a greater decrease in cell survival than luteolin alone, producing the most pronounced reduction in 4T1 cell viability among the tested conditions. Both luteolin and the exosomes exhibited a concentration-dependent effect, with cancer cell survival progressively declining as the dose increased (Figures 3 and 4).
Exosomes reduce tumor volume more effectively than luteolin: On day 1, there were no substantial differences in tumor volume between the treated and untreated groups (Figure 5A). After 14 days, tumor size increased markedly in the tumor group, whereas it decreased in the exosome-treated group. Tumor volume in the exosome group was reduced by approximately 57% compared with the tumor group on day 14, and by about 21% relative to its size on day 1. In contrast, the luteolin-treated group exhibited a 39% reduction in tumor volume relative to the tumor group. These findings indicate that exosome treatment was more effective in reducing tumor volume than luteolin treatment (Figure 5B).
Gene expression: Exosomes down-regulate RhoA gene expression: As illustrated in figure 6, RhoA gene expression was significantly elevated in the tumor group compared to the control group, highlighting its role in tumorigenesis. Treatment with luteolin led to a marked decrease in RhoA expression relative to the untreated tumor group, indicating luteolin’s capacity to suppress RhoA. Similarly, exosome treatment resulted in a substantial reduction in RhoA expression compared to the untreated group. Notably, 4T1-derived exosomes combined with luteolin exhibited an enhanced effect in downregulating RhoA gene expression (Figure 6).
Exosomes down-regulate ERK expression to a greater extent relative to luteolin: ERK gene expression was markedly elevated in the tumor group compared to the control group, indicating a significant association with tumorigenesis. Treatment with luteolin resulted in a notable reduction in ERK expression relative to the tumor group (Figure 7). However, exosome treatment induced a more pronounced and statistically significant decrease in ERK expression than luteolin alone, demonstrating greater efficacy in downregulating ERK gene expression (Figure 7).
Discussion :
Exosomes are cell-derived vesicles involved in various stages of cancer progression. They have gained recognition due to their remarkable stability in the bloodstream, non-toxicity, excellent biocompatibility, minimal immunogenic response, and capacity for specifically targeting cancer cells 15,16. In general, exosomes, contain a bunch of components including small interfering RNAs, microRNAs, membrane-associated proteins, and chemotherapeutic compounds, which have been shown to induce ameliorative effects for cancer treatment 12. Moreover, exosomes provide an optimal microenvironment that supports the effective activity of immunomodulatory factors. Indeed, altered-exosomes carrying bioactive molecules have been developed as novel tools capable of activating multiple stages of the cancer immunity cycle eliciting robust cancer-specific immune responses 12.
The emergence of novel bioactive compounds of natural origin, particularly those derived from plant sources, represents a promising and reliable therapeutic strategy for the treatment of various human cancers, owing to their selective action on specific molecular targets 17,18. Among these compounds, flavonoids such as luteolin are notable natural antioxidants that exhibit potent anticancer activity 19. Luteolin displays a broad spectrum of biological activities, including anti-inflammatory, anti-allergic, and anticancer effects, and function biochemically as either an antioxidant or a pro-oxidant. Moreover, these biological activities appear to be functionally interconnected; for instance, its anti-inflammatory effects may contribute to and reinforce its anticancer properties 20.
Luteolin has been shown to inhibit the proliferation of various tumor cell types in vitro and to effectively suppress tumor growth in vivo 21. During carcinogenesis, luteolin interferes with multiple stages of tumor progression—including cell transformation, metastasis, invasion, and angiogenesis—through diverse mechanisms such as kinase inhibition, cell cycle regulation, induction of apoptosis, and downregulation of transcription factors 22. Its anticancer effects are further supported by its ability to suppress the expression of oncogenic proteins, reduce tumor size and cell viability, decrease progestin-dependent VGF secretion, and enhance the expression of pro-apoptotic Bax. As a potent anticancer agent, luteolin demonstrates significant potential to inhibit proliferation and suppress the activation of key signaling molecules, including p-STAT3, p-EGFR, p-Akt, and p-Erk1/2, in EGF-stimulated MCF-7 breast cancer cells 23.
This study demonstrates that exosomes derived from luteolin-treated 4T1 cells can potentiate the anticancer effects of luteolin in breast cancer. Key outcomes, including reduced tumor size and modulation of cancer-associated markers, are likely attributable to specific alterations in the exosomal cargo. Exposure to luteolin may modify the gene expression profile of 4T1 cells, leading to the production of novel proteins, mRNAs, miRNAs, and other bioactive molecules, which are subsequently incorporated into secreted exosomes. These modified exosomes appear to suppress tumor growth more effectively than luteolin alone. In essence, the intrinsic therapeutic properties of exosomes, combined with their luteolin-induced cargo enrichment, may act synergistically or in combination with each other to enhance tumor inhibition.
Numerous studies have emphasized the synergistic potential of combining anticancer drugs with exosomes to enhance therapeutic efficacy. For instance, exosomes derived from Hepatocellular Carcinoma (HCC) cells were shown to improve the effectiveness of sorafenib in both in vitro and in vivo models. However, these exosomes also contributed to sorafenib resistance by activating the HGF/c-Met/Akt signaling pathway and inhibiting sorafenib-induced apoptosis 24. Similarly, exosomes derived from Natural Killer (NK) cells demonstrated cytotoxic effects against melanoma cells 25. With growing recognition of the diverse roles of exosomes, recent research has increasingly focused on their potential as a versatile drug delivery platform, capable of overcoming many limitations associated with synthetic nanoparticles 15.
Preclinical studies in animal tumor models have demonstrated that exosome-mediated delivery of chemotherapeutic agents can achieve superior antitumor effects compared with the administration of free drugs. For example, in a mouse model of colon adenocarcinoma, doxorubicin-loaded exosomes reduced tumor size more effectively than doxorubicin alone without detectable side effects 26. Similarly, Taxol-loaded Mesenchymal Stem Cell (MSC)-derived exosomes were used to treat metastatic breast cancer, resulting in a greater than 60% reduction in the weight of primary subcutaneous tumors following intravenous administration 27. Consistently, in the present study, exosome treatment led to approximately a 57% decrease in tumor volume, representing a more pronounced effect than luteolin treatment alone.
In the present study, exosomes induced a dose-dependent reduction in cell viability, achieving approximately a 70% decrease at a concentration of 320 μg/μl. Comparable findings have been reported in other studies. For instance, exosomes derived from placental stem cells selectively reduced the viability of prostate cancer cell lines without affecting non-cancerous human prostate cells 28. Likewise, MSC-derived exosomes were shown to inhibit the proliferation of Y-79 cells, as demonstrated by MTT assays, and to induce apoptosis in these cells within 72 hr 29.
Also, it was shown that exosomes from luteolin-treated cells exert these therapeutic effects by downregulation of the RhoA and ERK genes whereas the expression of both genes was elevated in 4T1 cells. RhoA is a crucial member of the Rho family of small GTPases, which regulate various cellular activities 30. Mutations and overexpression of Rho GTPases, in-cluding RhoA, have been observed in cancer cells. Dysregulated RhoA activity contributes substantially to carcinogenesis, making Rho GTPase signaling a potential target for cancer therapy 31. Studies have demonstrated that selectively inhibiting RhoA or its downstream effectors, such as ROCK, reduces proliferation and disrupts the G1-S transition in gastric cancer cell lines 32. The ERK pathway regulates cell proliferation, differentiation, survival, and motility. Disruption or aberrant activation of this pathway is commonly observed in multiple cancer types. When dysregulated, this pathway activates multiple transcription factors, promoting oncogenesis 33,34. Recently, ERK inhibitors have been developed 35. In one study, inhibiting the ERK/MAPK signaling pathway by employing an MEK inhibitor substantially reduced cell migration distance 36.
This study, like any others, suffers from certain limitations. The findings of this study should be confirmed in future in vivo studies with larger study groups and different concentrations of exosomes and luteolin. Also, possible existence of luteolin within the exosomes can be checked by certain methods like HPLC and LC-Mass. Exosome characterization would be better to perform using a panel of different markers. Also, it is better to assess the progression of tumor by more cancer markers and diverse methods.
Conclusion :
The research demonstrated that exosomes derived from luteolin-treated 4T1 cells effectively suppress the growth of breast cancer 4T1 cells, both in vitro by reducing cell viability and in vivo by decreasing tumor volume and downregulating the expression of tumor-associated genes, RhoA and ERK. The findings reveal potential of exosomes for augmenting anticancer effects of herbal-derived substances such as luteolin, which is advantageous for declining growing trend of breast cancer incidence throughout the globe.
Ethical approval: This study was approved by the regional ethics committee at Islamic Azad University under the specific ID number of IR.IAU.SRB.REC.1403.241.
Conflict of Interest :
The authors declared no conflict of interest.
Funding: Not applicable.

Figure 1. Characterization of exosomes derived from luteolin-treated 4T1 cells. A) Size distribution of exosomes determined by Dynamic Light Scattering (DLS). B) Representative Field Emission Scanning Electron Microscopy (FESEM) image showing the morphology of 4T1-derived exosomes.
|
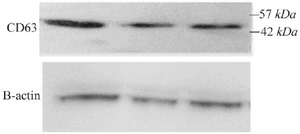
Figure 2. Western blot analysis of exosomes derived from luteolin-treated 4T1 cells. CD63 expression was detected, with β-actin serving as the loading control.
|
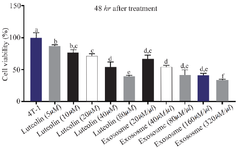
Figure 3. Effects of luteolin and exosomes derived from luteolin-treated 4T1 cells on cell viability. 4T1 cells were treated with increasing concentrations of luteolin and exosomes for 48 hr. Luteolin (80 μM) and exosomes (320 μg/μl) reduced cell viability by approximately 60% and 70%, respectively, in a dose-dependent manner. Exosome treatment showed greater cytotoxic efficacy compared to luteolin alone. Different letters indicate statistically significant differences between groups (p<0.05).
|
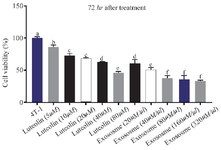
Figure 4. Effects of luteolin and exosomes derived from luteolin-treated 4T1 cells on cell viability after 72 hr of exposure. Treatment with luteolin (80 μM) and exosomes (80 μg/μl) resulted in a marked, concentration-dependent reduction in 4T1 cell viability. Exosome treatment demonstrated greater cytotoxic efficacy than luteolin alone. Different letters above the bars indicate statistically significant.
|
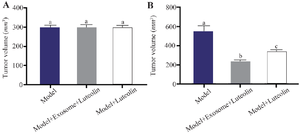
Figure 5. Effects of luteolin and exosome treatment on tumor volume. A) No significant differences in mean tumor volume were observed among groups on day 1. B) By day 14, tumor volume was reduced by 39% in the luteolin-treated group compared with the model group, whereas exosome treatment produced the greatest inhibition, resulting in a 57% reduction relative to the model group and a 21% decrease compared with baseline (day 1). Different letters above the bars indicate statistically significant differences between groups (p<0.05).
|
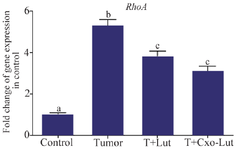
Figure 6. RhoA gene expression in control, tumor, luteolin-treated, and exosome-treated groups. Treatment with luteolin and exosomes derived from luteolin-treated 4T1 cells significantly downregulated RhoA expression compared with the tumor group. Different letters above the bars indicate statistically significant differences among groups (p<0.05).
|
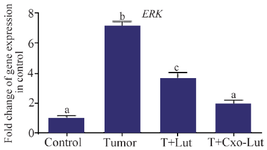
Figure 7. ERK gene expression in control, tumor, luteolin-treated, and exosome-treated groups. Luteolin treatment reduced ERK expression, whereas exosomes derived from luteolin-treated 4T1 cells elicited the most pronounced downregulation. Different letters above the bars denote statistically significant differences among groups (p<0.05).
|

Table 1. PCR primers for the assessed genes
|
|