Stress-Induced Proteomic Changes in the Hippocampus of Pregnant Wistar Rats
-
 M. Ardekani, Ali
Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran, Tel: +98 21 22432020 Fax: +98 21 22432021 E-mail: Ardekani@Avicenna.ac.ir
M. Ardekani, Ali
Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran, Tel: +98 21 22432020 Fax: +98 21 22432021 E-mail: Ardekani@Avicenna.ac.ir
-
Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Maghsudi, Nader
-
Neuroscience Research Center, Shahid Beheshti University of Medical Sciences, Tehran, Iran
-
Meyfour, Anna
-
Reproductive Biotechnology Research Center, Avicenna Research Institute, Tehran, Iran
-
Ghasemi, Rasool
-
Neuroscience Research Center, Shahid Beheshti University of Medical Sciences, Tehran, Iran
-
Lakpour, Niknam
-
Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Nooshinfar, Elahe
-
Department of Physiology, Faculty of Paramedical Sciences, Shahid Beheshti University of Medical Sciences, Tehran, Iran
-
Ghaempanah, Zahra
-
Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
Abstract: Stress is a threatening factor that all living organisms encounter throughout life. Depending on the type of stress, there are several mechanisms for keeping body homeostasis to minimize stress effects. Brain is an organ which shows high sensitivity to stress conditions. Although many studies have shown induced-stress effects on rat embryos, little is known about the mechanisms involved in coping with stress by female rats during pregnancy. In the present study, restraint stress method was applied because this technique has been widely used in animal models to induce both psychological and physical stress. Restraint stress was applied in regular sessions (1 and 3 hrs) in two groups of 6 pregnant Wistar rats and similar number of animals was used as control group receiving no stress. ACTH and corticosterone levels in plasma samples were shown to increase in response to stress treatments. On the last day of pregnancy, rat hippocampus from the brain of each animal in all three groups was removed and analyzed using 2 Dimensional Gel Electrophoresis (2DE) technique. Using Image Master Software, approximately 2000 proteins were detected in the 2D gels analyzed, among which 34 proteins exhibited differential expression. These results indicate that the proteome patterns from the hippocampus of pregnant rats subjected to 1 and 3 hr of stress differs significantly from the control (unstressed) group. Future mass spectrometry identification of the 34 protein spots discovered in this study should allow a more precise understanding of molecules and cellular pathways involved in stress-induced responses during pregnancy.
Introduction :
Stress threatens living organisms throughout their life. This is the way body responds to something outside the normal (i.e. risky, unknown or disturbing) (1). Cohen et al have offered a helpful meaning of stress: ‘‘when environmental demands [internal or external; real or imagined] tax or exceed the adaptive capacity of an organism … psychological and biological changes that may place persons at risk for disease… follow” (2). Depending on the type of stress, there are several mechanisms for keeping body homeostasis to minimize stress effects. It has been suggested that stress is ‘anything, that induces increased secretion of glucorticoids’ (3). Glucocorticoid hormones contribute in normal maturation of the developing axons and dendrites, and the cell survival (4).
When a person encounters stressors that are demanding and/or threatening, two principal physiologic cascades occur in the brain. One involves the autonomic nervous system and the release of catecholamines, specifically norepinephrine and epinephrine and the second involves the Hypothalamic-Pituitary-Adrenal (HPA) axis, Corticotropin-Releasing Hormone (CRH), Adrenocorticotropin hormone (ACTH), and cortisol (5). Therefore, Central Nervous System (CNS) has an important role in the response to physiological stress.
Hippocampus is a brain region that has protracted development, retains the ability to produce new neurons in adulthood in most mammalian species (6) and plays an important role in memory and executive functions (7). High density of glucocorticoid receptors has been reported in the hippocampus and has been shown to be the potential targets of corticosteroid hormone and vulnerable to the effects of stress (8). High levels of stress may lead to permanent damage to certain functions of the developed adult brain (9) and hippocampal volume and neurogenesis have been shown to decrease in depressed patients (10). Significantly, it has been demonstrated (11) that chronic stress and high amount of corticosterone suppress hippocampal neurogenesis in both male and female rats. Interestingly, females have been reported to respond differently toward stress or elevated glucocorticoids when compared to males (12). Females have been also shown to be more susceptible to stress condition and the rates of occurrence for depression in women are reported to be twice as men (13).
Many studies have been performed to understand the effects of stress hormones during different phases of reproduction and how they affect the brain and behavior of fetus. Such investigations have identified relationships between ‘‘stress’’ hormones (i.e., ACTH and cortisol) and either an increased occurrence of Preterm Birth (PTB) and/or increased maternal blood levels of CRH in pregnancy (14). In an experiment by Wadhwa et al (15), a correlation between elevated maternal plasma CRH levels at 33 weeks of gestation, and a 3.3-fold increase for spontaneous PTB was reported. Furthermore, maternal stress and increased levels of glucocorticoids in utero have been demonstrated to have extensive acute effects on neuronal structure causing morphological changes in the hippocampus and other brain structures of the embryo (some of which may not become obvious until adulthood). For instance early exposure to stress makes hippocampal atrophy, reduction in dendritic branching and alters the development of neural connection and neurotransmitter systems in the developing brain which may contribute to neurodevelopmental disorders and psychiatric diseases in later life (8,16).
Although numerous studies have been reported on sex differences in response to stress and effects of stress on the brain development in rat embryos, little research has focused on the question of how stress or elevated glucocorticoids during pregnancy or postpartum can affect the brain in the pregnant female rats (17). Recently, proteomics approach has been applied by Skynner H.A. et al (18) to investigate the proteomic changes occurring in mouse brain following chronic exposure to corticosterone using two-dimensional Differential In-Gel Electrophoresis (DIGE). In another study, molecular changes in protein levels of rat hippocampus caused by exposure to repeated psychosocial stress was evaluated by 2 DE and results showed that the pattern of protein alterations induced by repeated stress was quantitatively and qualitatively different from that observed after a single exposure (19).
To better understand the molecular pathways and the role of molecules involved in stress-induced mediated brain effects in the hippocampus region, we studied the protein profile of hippocampus in the female rats subjected to restrain stress during the last week of pregnancy, using 2 DE proteomics approach.
Materials and Methods :
Animal
Female Wistar rats 180-220 g, randomly assigned into three groups (each group of 6 rats) were housed in a temperature 22-25C and humidity-controlled room. Animals were maintained under a 12:12 hr light/dark cycle. Food and water provided ad libitum except for stress sessions. Animals were housed with male rats for one night for mating and the day after that was assumed as the first day of pregnancy. On the 14th day, based on weight gains in pregnant rats, they were separated and housed in a new cage and between 14th to 20th day of pregnancy, rats were exposed to daily restrain stress for 1 hr (1 hour group) or 3 hr (3 hours group); control group did not receive stress.
Immobilization stress
To induce stress, animals were immobilized in a plastic rodent restrainer, adjustable to animal size so that animal's movement was completely restricted. Stress sessions were started at 9 AM and after each stress session, rats were returned to their respective cages. On the last day of pregnancy, rats were lightly anesthetized with CO2, decapitated and their hippocampus were dissected out and immediately frozen in liquid nitrogen and stored at -80 C for later analysis.
Hormone and statistical analyses
To measure plasma levels of ACTH and corticosterone hormones, ELISA tests were performed as directed by the manufacturers. The kit for ACTH assay was obtained from Phoenix Pharmaceuticals Inc. Burlinngame, USA and the kit for Corticosterone was obtained from DRG instruments GmbH, Marburg, Germany. Statistical analysis was performed on the changes in plasma levels of ACTH/corticosterone using one-way ANOVA followed by Tukey's post hoc test. Data are presented as mean ± S.E.M. from three separate groups of six animals (total number of 18 animals).
Sample preparation and two dimensional gel electrophoresis (2DE)
Hippocampus were homogenized by pestle in lysis buffer containing 7 M Urea, 2 M Thiourea, 4% CHAPS(3- (3-Cholamidopropyl) dimethylammonio)-1-Propanesulfonic Acid), 20 mM Tris, 10 mM DTT (Dithiothreitol), 1 mM PMSF (Phenylmethanesulfonylfluoride), 1 mM EDTA (Ethylenediaminetetraacetic acid), and Protease Inhibitor (one tablet in 2 ml lysis buffer) (Roche). Homogenates were sonicated five times on ice for 30 s and left for one hr at room temperature. Lysates were centrifuged at 14000×g for 60 min at 12 C. Protein concentrations were determined by Bradford assay. One hundred and twenty µg from each sample was resuspended in rehydration buffer containing 8 M urea, 4% CHAPS, 2 mM TBP (tributyl phosphate), 0.2% Ampholyte, 10 mM DTT for 16 hr and then loaded onto 17 cm immobilized (pH=3-10) nonlinear gradient strips (Bio-Rad, Hercules, CA, USA).
Strips were focused at 20 C with the following program: 0-250 V for 20 min with linear increase, followed by linear increase to 10000 V, and remained on 10,000 V to achieve total 50,000 V h in a PROTEAN® i12TM IEF Cell (Bio-Rad). The strips were reduced in equilibration buffer containing 20% glycerol, 2% SDS (Sodium Dodecyl Sulfate), 6 M urea, 50 mM Tris-HCl and 2% DTT for 20 min and subsequently alkylated in the same buffer containing 2.5% Iodoacetamide instead of DTT for 20 min. The IPG (immobilized pH gradient) strips were placed on 12% Polyacrylamide gels and electrophoresed initially for 30 min at 16 mA/gel and then 5 hr at 24 mA/gel using the protein Xi-II cell (Bio-Rad laboratories). Resulting gels were stained with silver nitrate (0.2%).
Image analysis
Silver stained gels were scanned by
Densitometer GS-800 (BioRad) and subsequently were analyzed by the ImageMasterTM 2D platinum 6.0 software (Amersham Biosciences). Spot detection and matching were performed and volumes of protein spots were appraised and matched among gels. Data obtained from 2DE gels of 1 hr stress-induced samples and 3 hr stress-induced samples (prepared from three repeats) were compared with the control group using Student’s t-test on %vol of matched spots showing greater than 1.5 fold change in expression levels.
Results :
Stress effects on corticosterone and ACTH changes
In this study we applied restraint stress to pregnant Wistar rats in the 3rd week of pregnancy (days 14th to 21th) and measured stress associated hormones. Results from hormone analysis confirmed stress induction and showed significant alterations in plasma corticosterone and ACTH levels in 1 and 3 hr stress-induced groups (Figures 1 and 2). Amount of corticosterone in maternal plasma was increased in 1 hr by 2.4 and in 3 hr by 1.7 folds, respectively (Figure 1). As shown in figure 2, ACTH levels were increased in 1 hr by 1.6 and in 3 hr by 1.5 folds, respectively. The data from both groups receiving stress were compared with control group which received no stress.
Proteomic changes of hippocampus proteins following 1 hr stress induction
2DE was performed on proteins extracted from hippocampus of control and 1 hr stress-induced rats using 17 cm IPG strips with nonlinear pH (range 3-10) (Figures 3A and 3B). Normalized volumes (%vol) of the matched spots (the total number of spots were 2,267) in replicates of control and 1 hr stress-induced samples were analyzed by student’s t-test. Twenty five spots were found to be significantly different in %vol (at least 1.5- fold change) when compared with the control group. As shown in table 1, 11 protein spots were found to be up-regulated and 14 protein spots exhibited a decreased expression. The range of protein spot sizes vary between 6-100 kDa indicating a differential protein expression in hippocampus from 1 hr stress-induced pregnant rats as shown in figure 3B.
Proteomic changes of hippocampus proteins following 3 h stress induction
In figure 3C, the proteomic changes in hippocampus of 3 hr stressed rats are shown. The total of 2476 spots were detected and 9 protein spots demonstrated a significant percent volume changes as shown in table 2. Among the 9 spots showing difference in protein expressions (control versus 3 hr stress-induced samples), five spots were found over-expressed, three spots were found under-expressed and one protein spot disappeared in 3 hr stress-induced samples. The range of protein spot sizes vary between 20-120 kDa indicating a differential protein expression in hippocampus from 3 hr stress-induced pregnant rats as shown in figure 3C.
Discussion :
This was the first study to investigate the proteomic changes occurring in hippocampus of pregnant rats subjected to repeated restraint-stress. The proteomic map we obtained in this study was very similar to the protein map of rat hippocampus reported previously (20). However, the 2DE gels in our study appeared to have a higher resolution, less streaking and more protein spots (Figure 3). In this study we developed a pregnant rat model to investigate the proteome of hippocampus subjected to restraint stress. The reason the stress effects on the proteome of hippocampus was investigated in this study was due to the role that hippocampus plays in learning and memory and its susceptibility to stress and senescence (21) and its potential role in neurodegenerative diseases and some psychiatric disorders (22).
Activation of the HPA axis is a major response of mammalian brain to stress hormones. As we have shown in this study (Figures 1 and 2), the chronic stress for 1 and 3 hr lead to activation of HPA as represented by ACTH and corticosterone release. The increase in plasma corticosterone and ACTH levels in stress-induced groups relative to the control group (Figures 1 and 2) demonstrated that the rats were under actual stress and all measurements made were representative of the stress-induced state. As it can be seen in figure 1, corticosterone levels increase in 1 hr and decrease in 3 hr. In parallel ACTH levels (Figure 2) increase in 1 hr and decreases in 3 hr, respectively. Both ACTH and corticosterone are known to mediate adaptation to stress and regulate termination of the stress response through negative feedback mechanism at the level of the HPA axis (23). In the brain, the Glucocorticoid Receptor (GR) modulates many neuronal activities, including stress-related activation of the HPA axis (24). It is reported that GR in the hippocampus play a major role in the adaptation to chronic stress (25) and its expression at the mRNA levels during repeated restraint stress is potentially associated with activation of the HPA axis (26). We suspect that adaptation by the animals to restraint stress leads to lower production of corticosterone and thus lower production of ACTH in 3 hr.
Protein levels of rat hippocampus exposed to repeated stress has been reported and changes in the pattern of proteins were demonstrated to be different from that observed after a single exposure (19). In this study, as the pattern of proteome changes in the hippocampus of rats subjected to 1 hr and 3 hr repeated stress were compared, a significant difference was observed (Figures 3B and 3C). The intensities of approximately 2000 protein spots were analyzed in both 1 hr and 3 hr stress-induced groups and 34 spots were found to be significantly changed in the hippocampus by different stress treatments. It is interesting that the differential expression of more than 70% of protein spots (25 spots) were found to be changed in the 1 hr stress-induced samples and 9 spots in the 3 hr stress-induced samples. These results indicate that the proteome pattern in the hippocampus of pregnant rats vary when subjected to different amount of stress. Furthermore, it appears that with increasing stress time, the number of protein spots decline from 25 to 9, perhaps due to the adaptation mechanisms that the brain employs during the time of stress. Although we have not been able to sequence the protein spots in this study due to certain limitations, the molecular weight markers indicate that almost all protein spots were localized in the 10-120 kDa range (Figures 3B and 3C) and most changes were detected in the 20-120 kDa range.
Part of the brain's response to stress is changes in the expression of neutrophins such as Brain Derived Neurotrophic Factor (BDNF), nerve growth factor and their protein kinase receptors of the trk family (27). BDNF is the most abundant and extensively expressed neutrophin, a family of structurally related proteins required for the development and function of the vertebrate nervous system (28). It has been demonstrated that BDNF may be involved in stress-induced adaptation in adult (29). Studies have shown that the levels of BDNF protein and mRNA were increased in the pituitary glands of rats stressed for 60 min while its expression level were decreased following stress for 180 or 300 min (30). Also the increased expressions of BDNF mRNA and protein were observed in the hippocampus of rats which had been treated with short-term sleep-disturbance (31). Parallel to these studies, much research have shown that single immobilization stress for 8 hr (longer than 2 hr) (32) or repeated immobilization stress (2 hr/day for 7 days ) (27) decrease BDNF mRNA throughout the hippocampus, as measured by in-situ hybridization. Due to the critical role of hippocampus in memory formation in human and possibility of similar role in rats (21), shorter times of exposure to stress can directly or indirectly cause an increase in neutrophins, which may contribute to store the information of this event. It is thought that maintaining such information in the hippocampus can help the animal to have an appropriate response to new stressors in the future.
Moghadam et al (33) revealed twenty minutes restraint-stress increases the neuronal release of glutamate and aspartate in hippocampus, prefrontal cortex and nucleus accumbens, but it was shown that extracellular glutamate concentrations in the hippocampus returned to baseline by the end of a 1 hr restraint-stress (34). Probably glutamate and other neurotransmitters are involved in the regulation of BDNF expression during stress and some of them could be responsible for decreased level of BDNF at longer exposure time to stress (32). The results of these studies may suggest that the extensive proteomic changes we have observed in the hippocampus of pregnant rats subjected to repeated 1 hr stress and the limited alterations in the proteomic expression profile of the rats subjected to 3 hr stress, may be indicative of different levels of expressional changes in neurotransmitters, neutrophins, BDNF and other unknown proteins associated with memory creation. We speculate that the significantly lower alterations in the proteome of rat hippocampus subjected to longer time of stress (3 hr, Figure 3C) may be due an adaptation of brains of these animals to stress in order to prevent harmful effects of stress to themselves and/or the embryo.
Very few proteomics studies have been reported on the effects of stress on hippocampus in the mammalian system. It is known that following psychosocial stress in rat, protein changes occur in the hippocampus region and these proteins belong to different categories of cellular functions including: protein folding, signal transduction, synaptic plasticity, cytoskeleton regulation and energy metabolism (19). In another study, DIGE technique was used to identify 150 corticosterone-induced protein spot changes in the rat hippocampus and connected brain regions (18). Recently, a proteomics study on the whole brain homogenate of an anxiety mouse model revealed extensive protein changes involved in either serotonin receptor protein packets, metabolic events, oxidative, synaptic docking or signal transduction pathways (35).
Conclusion :
In conclusion, this study for the first time demonstrates that 2DE is a useful method for studying the proteomic changes in the hippocampus of rats subjected to stress during pregnancy. Our data clearly demonstrates that exposure to repeated cycles of stress (as little as 1 hr everyday for 7 days) during pregnancy can result in significant differential protein expressions. The future identification of protein spots discovered in this study by mass spectrometry techniques should allow a more detailed analysis of pathways and molecular mechanisms involved in the stress-induced changes in the proteome of hippocampus from rat brains during pregnancy. Such studies should further our understanding of female mammalian adaptation systems to stress during pregnancy and the built-in mechanisms in the mammalian brain which helps the survival of the embryos during stressful times. Specific drug-targeting treatment protocols can be envisaged for management of stress if similar pathways and mechanisms are found in humans in the future.
Acknowledgement :
This study was supported by a grant from the Neuroscience Research Center, Shahid Beheshti University of Medical Sciences.
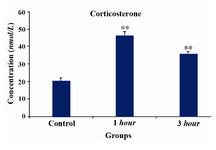
Figure 1. Measurement of corticosterone hormone in three groups of pregnant rats (control-no stress, 1 hr and 3 hr stress-induced rats). Concentrations are expressed as mean ± S.E.M. (as described under Materials and Methods); n=3 animals per experimental group. Asterisks indicate significant differences between treated groups. Statistical analysis was performed using one-way ANOVA followed by the Tukey’s post hoc test (*p<0.001, 1 hr stress versus control; *p<0.001, 3 hr stress versus control)
|
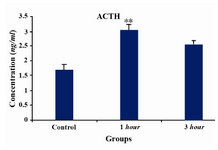
Figure 2. Measurement of ACTH in three groups of pregnant rats (control-no stress, and 1 hr and 3 hr stress-induced rats). Concentrations are expressed as mean ± S.E.M. (as described under Materials and Methods); n=3 animals per experimental group. Asterisks indicate significant differences between treated groups. Statistical analysis was performed using one-way ANOVA fol-lowed by the Tukey’s post hoc test (**p<0.001, 1 hr stress versus control; *p<0.01, 3 hr stress versus control)
|

Figure 3. A representative image of silver nitrate-stained 2DE gel from control (A), 1 hr (B), and 3 hr (C) stress-induced hippocampus rats. Proteins were separated on a pH 3-10 NL IPG strips in the first dimension and on 12% SDS polyacrylamide gel in second dimension
|
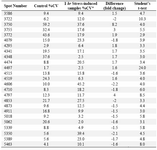
Table 1. Volume changes for 2DE spots reflecting protein expression changes in hippocampus of 1 hr stress-induced rats compared with the control group
٭CV: The coefficient of variation (Coef. Variation) is the dispersion divided by the central tendency. It measures the relative variability of the spots in a match by correcting for the magnitude of the data values, thus giving a measure that has no units. The critical value for p = 0.05 is 2.776. The numbers reported in this table are values above 2.77.
|
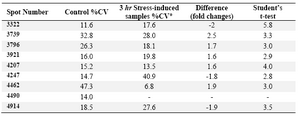
Table 2. Volume changes for 2DE spots reflecting protein expression changes in hippocampus of 3 3 hr stress-induced rats compared with the control group
٭CV: The coefficient of variation (Coef. Variation) is the dispersion divided by the central tendency. It measures the relative variability of the spots in a match by correcting for the magnitude of the data values, thus giving a measure that has no units. The critical value for p = 0.05 is 2.776. The numbers reported in this table are values above 2.77.
|
|