Transient Co-Expression of Bioactive Murine Interferon-Gamma and HBsAg in Tobacco and Lettuce Leaves
-
Mohammadzadeh, Sara
-
Department of Medical Biotechnology, Faculty of Medicine, Kermanshah University of Medical Sciences, Kermanshah, Iran
-
Amiri, Mahshid
-
Laboratory of Cellular and Developmental Biology, NIDDK, National Institutes of Health, Bethesda, United States of America
-
 Ehsani, Parastoo
Department of Molecular Biology, Pasteur Institute of Iran, Tehran, Iran, Tel: +98 21 64112219 Fax: +98 21 64112803; E-mail: p_ehsani@yahoo.com
Ehsani, Parastoo
Department of Molecular Biology, Pasteur Institute of Iran, Tehran, Iran, Tel: +98 21 64112219 Fax: +98 21 64112803; E-mail: p_ehsani@yahoo.com
Abstract: Background: The synchronous expression of antigen and adjuvant proteins in plant hosts presents an intriguing potential for vaccine production and the enhancement of appropriate immune responses. In this study, we examined the expression of bioactive murine interferon-gamma (mIFN-γ) along with HBsAg in tobacco and lettuce leaves aimed to further perform the analysis of immune responses in the mouse model.
Methods: Monocistronic and bicistronic cassettes, carrying genes encoding mIFN-γ and HBsAg in various orders, were constructed. These cassettes were placed under the control of the 35S CaMV promoter and included the 5ʹ leader sequence of Tobacco Ech Virus (TEV). Through Agrobacterium infiltration, the cassettes were transferred into plant leaves. The concentration of mIFN-γ in different constructs and HBsAg was tested by ELISA. Murine IFN-γ was characterized through Western blotting, and its bioactivity was evaluated by assessing the up-regulation of MHC class II in macrophages derived from mouse bone marrow.
Results: Extracts of agroinfiltrated leaves contained recombinant mIFN-γ and HBsAg proteins at about 14 unit/mg and 50 ng/mg of soluble protein, respectively. Subsequently, mIFN-γ was purified from the plant extract and its ability to up-regulate MHC class II in mouse bone marrow-derived macrophages was confirmed by immunofluorescence.
Conclusion: The co-expression of recombinant HBsAg and mIFN-γ using TEV 5ʹ leader-based cassettes in tobacco and lettuce leaves produced both proteins with active mIFN-γ in different concentrations. The attractive utility and feasibility of using plant transient co-expression systems aimed to co-delivery of vaccine antigen and appropriate cytokine to elicit immune response for different applications.
Introduction :
Hepatitis B virus (HBV) infection with an estimated 296 million chronic carriers remains a significant global health problem 1. Recombinant HBsAg vaccines with alum-adjuvant, like Recombivax HB and Engerix, are widely used in preventing and controlling strategies in the world. However, additional strategies are introduced to improve the immune response and seroconversion rates in vaccine recipients, especially in immune-compromised individuals 2. Therefore, to enhance the immune response to vaccines, cytokines have been considered as adjuvant for their efficiency and safety 3.
Interferon-gamma (IFN-γ) is a potent immunostimulator with 100–10,000 times more active compared to other types of interferons including IFN-α and IFN-β 4. It is mainly produced by activated T-lymphocytes and Natural Killer (NK) cells, mediating T cell proliferation and differentiation to polarize Th1 and cytotoxic immune response 5. It also affects macrophage activation and augmentation of the MHC class I and II molecules 6. The important role of this cytokine in evaluating Th1 immune response, antiviral activity, and tumor reduction effect leads to extensive application against the intracellular pathogens and various tumors 7.
Fast and continuous protection through the co-administration of a vaccine and IFN-γ as antiviral agents has been demonstrated for several diseases, including foot-and-mouth disease 8. Furthermore, some pre-clinical and human studies proposed that adjunctive immunotherapy with IFN-γ could promote establishing protective immunity against pathogens including mycobacteria 9 and HIV 10. IFN-γ as an adjuvant enhanced specific Abs response rate to hepatitis B vaccination of hemodialysis patients 11. Additionally, the results suggested that co-immunization of mice with plasmid DNA encoding mouse IFN-γ enhanced an IgG2-biased humoral response to HBV DNA-based vaccines 12.
However, IFN-γ demonstrated species-specific differences in binding properties to its receptors, mostly non-existent or minimal cross-reactivity 13. Therefore, in vaccine research, the production of heterologous interferon with species origin is needed to evaluate nonspecific immunity and modulation of the adaptive immunity in the model animals 14.
A range of expression systems has been utilized for the production of recombinant IFN-γ 15. In the past two decades, plant-based production systems have attracted attention due to low production costs and quick scale production and are also considered for oral delivery of several recombinant proteins particularly vaccine antigens without or with minimal formulation 16. In this regard, since orally administered IFN maintains biological effects in humans and other animals 17, plant-produced IFN could be directly consumed without expensive processes of purification. The co-expression of IFN-γ and Mycobacterium tuberculosis antigens in transgenic plants has been patented for use as an edible vaccine 18.
Plant-based systems were successfully used for the production of avian 19 and bovine IFN-γ 20. The human IFN-γ was also synthesized in different plant expression systems such as in tobacco 21, rice cell suspension culture 22, transgenic brassica 23, and tomato 24. A considerable expression level comprising 6% of the soluble protein content was achieved in transplastomic tobacco plants 25.
However, the mouse and human IFN-γ share only about 40% sequence homology and are species-specific; therefore, reliance on the cross-species tests might be controversial for immunological studies in mouse models. Here we report the efficient expression of biologically active plant-based mouse IFN-γ aimed to apply in preclinical tests performed in murine models. To our knowledge, the plant-based murine IFN-γ has not been produced and the bioactivity has not been characterized yet in other studies.
We used the expression cassette carrying 35S CaMV promoter followed by a Tobacco Etch Virus (TEV) 5ʹ leader sequence. It is shown that the TEV 5΄Untranslated Region (UTR) sequence promotes mRNA translation using a cap-independent pathway 26. Moreover, when positioned internally within a bicistronic construct, it also promotes translation of the second ORF due to Internal Ribosome Entry Site (IRES) activity 27. Therefore, the co-expression of both free antigen and immune stimulatory proteins in the plant cell increases the data on applying monocistronic and bicistronic cassettes containing TEV leader sequences.
Materials and Methods :
Plasmid construction: Murine IFN-γ (mIFNG) gene was PCR amplified from the pIFNGM vector (kindly provided by Pierre Marie Curie, Institute Paris, France). The pCMV-S2S plasmid was the source of HBsAg gene (a gift from Dr. ML Michel, Pasteur Institute, Paris, France). Dr. H Ofoghi (IROST, Tehran, Iran) has provided pRTL plasmid 28 containing CaMV 35S dual promoter-enhancer and TEV 5ʹ leader sequence. The above vector was used to produce monocistronic and bicistronic constructs by inserting different genes in XhoI and EcoRI sites for upstream and BamHI site for downstream of the TEV leader sequence. mIFNG gene was amplified using forward and reverse primers with the following sequences; 5ʹ-AGCATATGCCTCGAGGATCCATGAACGCTACACACTGC-3ʹand 5ʹ-TATACTATTCTCGAGGATCCTCAGCAGCGACTCCTTT TCCG-3ʹ, respectively. These primers contain sites for the restriction endonuclease XhoI (underlined) and BamHI (italic). The gel-purified 508-bp PCR product was excised with either XhoI for cloning upstream of TEV leader or BamHI for cloning downstream of the TEV leader into the pRTL (Figure 1).
The gene encoding HBsAg was PCR-amplified using forward primer; 5ʹ-TTTTATAGATCTGAATTCAT CTTCTCGAGGATTGGGGAC-3ʹ and reverse primer; 5ʹ-TTTTTGGAATTCAGATCTAGAGTAACCCCATC TC-3ʹ which comprise sites for the restriction endonuclease BglII (underlined) and EcoRI (italic) sites. The amplified 783-bp PCR product was digested using either EcoRI for cloning upstream of TEV leader or BglII for cloning downstream of the TEV leader into the corresponding sites of pRTL in two configurations as shown in figure 1. Amplification was performed at 94°C for 5 min followed by 30 cycles (1 min at 94°C, 30 s at 58°C, 1 min at 72°C) and 7 min at 72°C. The molecular manipulations were carried out according to Sambrook et al 29. The sequencing was done by MWG Biotech AG (Ebersberg, Germany) and confirmed by ClustalW multiples alignments program against EMBL gene bank sequence for mouse ifng gene.
The Sure electroporation-competent Escherichia coli (Stratagen, USA) was used for transformation by cloned pRTLs vectors containing the expression cassettes. The expression cassettes were digested by HindIII, and sub-cloned into the binary vector pDE1001 (kindly supplied by Dr. Ann Depicker, Ghent University, Belgium) 30 as shown in figure 1.
The recombinant pDE1001 vectors were transferred into electrocompetent Agrobacterium tumefaciens C58CI RifR (a gift from Dr. G De Jaeger, Ghent University, Belgium) and cultivated on YEB medium supplemented with Kanamycin (50 µg/ml), Spectinomycin (50 µg/ml) and Rifampicin (100 µg/ml) at 28°C for 48 hr.
Agroinfiltration of leaves: The leaves of tobacco (Nicotiana tabacum L. cultivar Xanthi) and lettuce (Lactuca sativa L.) were agroinfiltrated 31. Briefly, 50 mL of fresh YEB media containing 10 mM 2-(n-morpholino) ethanesulfonic acid (MES) (Merck, Germany) pH=5.6, 20 µM acetosyringone (Sigma, USA), and the antibiotics were inoculated with 48 hr grown bacterial cultures of Agrobacteria carrying different vectors then the overnight cultures were grown to log phase (OD600=0.8-1.0) and harvested by centrifugation at 5000 rpm for 10 min at 4°C. The precipitated cells were suspended in media containing Murashige and Skoog salts, 10 mM MES, 20 g/L sucrose, pH=5.6, and 200 µM acetosyringone (MMA medium) up to a final OD600 2.4 and kept at 22°C for 1 hr. Then leaves were put in Agrobacterial suspension and vacuumed at 0.1-1 mbar twice, each time for 10-15 min. The infiltrated leaves were washed and kept on filter paper in petri dishes and placed at 22°C with 16 hr light/8 hr dark condition for 3-5 days.
Protein extraction of agro-infiltrated leaf: For the protein extraction, the leaves were ground to a fine powder in liquid nitrogen and mixed with one volume of cold extraction buffer (20 mM sodium phosphate; pH=7.5, 1% Triton X-100, 0.15 M NaCl, and 2 mM PMSF). The insoluble material was removed following centrifugation at 17000 g for 15 min. The supernatants were dispensed in tubes and stored at -80°C. Bradford protein assay was used for the determination of Total Soluble Protein (TSP) concentration.
Determination of expression of HBsAg: The transient expression of HBsAg in agroinfiltrated leaves was evaluated using the Monolisa HBsAg ELISA Kit (BioRad, Irvine, CA, USA) following the manufacturer's instructions. Additionally, a sandwich ELISA assay was developed to further investigate the expression. Briefly, in the second assay, the wells were coated with 2 µg/ml monoclonal anti-HBsAg antibody 112A26 and incubated overnight at 4°C. Subsequently, the wells were blocked with 1% gelatin for 2 hr at 37°C. 50 μg of TSP from each sample were added to the coated wells in duplicates and incubated for 1 hr at 37°C. Rabbit L3a_S2S antibody (kindly provided by Dr. Marie-Louise Michel from the Pasteur Institute of Paris) was used as the secondary antibody and incubated for 1 hr at 37°C. All incubations were followed by washing three times with PBST (PBS containing 0.1% Tween 20). Finally, 100 µL/well of a diluted (1/3000) peroxidase-labeled goat anti-rabbit IgG (H + L) antibody (Vector Laboratories, Burlingame, CA, USA) was added and incubated for 1 hr at 37°C. After washing, the enzymatic reaction developed with TMB substrate and measured at 450 nm.
IFN-γ expression assays: For western blotting, samples were loaded on 12% SDS PAGE and following electrophoresis transferred to the nitrocellulose membrane. After blotting, the nitrocellulose paper was incubated in 3% BSA overnight at 4°C. Subsequently, the paper was treated with a 0.5 µg/ml biotinylated anti IFN-γ antibody (Pharmingen, San Diego, USA) for 1 hr at 37°C. After washing, the paper was then exposed to streptavidin-conjugated peroxidase (diluted 1:15,000) at room temperature for 1 hr. The immunoblot was visualized using ECL Plus detection reagents (Amersham Pharmacia Biotech, Little Chalfont, UK) and developed on autoradiography film (HyperfilmTM ECL).
A sandwich ELISA assay for evaluating the transient expression of IFN-γ was done according to de Moraes 32. Briefly, ELISA plates were coated using 1 µg/ml mouse anti-IFNγ antibody (PharMingen, San Diego, USA). Then they were blocked using 1% gelatin in TPBS, following washes the plates were incubated with 100 µl of leaf extracts for 90 min at room temperature. The process was continued with 3 washes with TPBS, adding 0.5 µg/ml biotinylated anti-IFNγ and further incubation for 90 min at room temperature. Following three washes, streptavidin-peroxidase was added to plates and incubated for 1 hr. After further washings, OPD (1,2 phenylenediamine dihydrochoride) (DAKO) was added as substrate, and absorbance was read at 492 nm.
Immunoaffinity purification of IFN-γ: The affinity column was prepared using the Seize™ X Protein G Immunoprecipitation kit (Pierce, Rockford, IL, USA) according to the protocol recommended by the company. Briefly 400 µg of the primary antibody (Pharmingen) was coupled to the 0.4 ml of immobilized protein G. Then it was cross-linked by 2 mg of Disuccinimidyl Suberate (DSS). The prepared column was loaded with 500 µl of the plant extract. Elution was done by immuno-pure elution buffer pH=2.5-3 which was neutralized by 1 M Tris solution pH=9.5 while it was eluted from the column in 10% final concentration.
Biological activity analysis of IFN-γ: BALB/c bone marrow-derived macrophages were harvested and cultured for 5 days according to Antoine et al 33. Then cells were washed and cultured for 18 hr in the presence of 50 units of murine recombinant IFN-γ (rIFNγ) that was kindly provided by Dr. G.R. Adolf (Emst Boehringer Institute Fur Arzneimittelforschung, Vienna, Austria), column purified plant-derived mIFNγ and negative control. For detection of MHC class II up-regulation, the cells were used for immune-fluorescence staining. After fixation in paraformaldehyde (4%), they were incubated for 1 hr at room temperature with 1 µg/ml anti–MHC II (clone M5/114). Then cells were washed and incubated for 1 hr at room temperature with 10 µg/ml of donkey anti-rat IgG (Fab) fragment conjugated to Fluorescein Isothiocyanate (FITC) (Jackson ImmunoResearch company).
Statistical analyses: Prism 5.0 software (GraphPad, USA) was used for the data handling and results are presented as means ± standard error of the mean (SEM). statistical analysis was carried out using one-way ANOVA and non-parametric tests. Statistical significance was set at p≤0.05.
Results :
Construction of expression cassettes: mIFNG and HBsAg genes were amplified and digested by the restriction enzymes and ligated into pRTL in the presence of noncoding TEV 5ʹ leader sequence, the elements of the plant expression cassette used for the co-production of mouse IFN-γ and HBsAg were shown in figure 1. The sequenced mIFNG fragment was identical with the coding region (110-577) of Mus musculus interferon gamma (mIfng), mRNA (NCBI RefSeq: NM_008337.4) and the corresponding amino acid sequence, and its post-translational modification is presented in figure 2.
Characterization of recombinant mIFN-γ produced in plant: Western blotting analysis confirmed the presence of mIFN-γ in plant extract (Figure 3). Western blot analysis of plant extract after treating the purified IFN-γ with 0.1% SDS for 1 hr at 37°C, showed the monomer protein which weighted about 25 and enriched about 60-70 kDa (Figure 3).
Bicistronic protein expression analysis: ELISA analyses were performed to detect and quantify the expression of HBsAg and mIFN-γ. Both HBsAg and mIFN-γ proteins were detected in tobacco and lettuce leaf extract. The HBsAg transient expression level was varying from 15-50 ng/mg of total soluble protein from leaf extract. The results of the ELISA test indicated that the most expression level of mIFN-γ is about 14 units/mg of total soluble protein. The comparison analysis of different expression cassettes by ELISA confirmed that the expression level of mIFN-γ was several folds higher in tobacco leaves agroinfiltrated with IFN-γ-TEV-HBsAg bicistronic construct. Although no statistically significant differences were observed in the results (Figure 4).
Bioactivity of IFN-γ: The assay of the bioactivity of IFN-γ relied on the upregulation of MHC class II molecules on mouse bone marrow derived monocytes. For testing the bioactivity of IFN-γ, an affinity column purified IFN-γ was used because high concentration of Agrobacterium LPS can provoke down-regulation of monocyte MHC class II surface expression 34. As it is shown in figure 5, the macrophages treated with plant-derived purified mIFN-γ have generated the MHC Class II on the monocytes in the presence of an L-conditioned medium.
Discussion :
Despite some reports on plant produced human IFN-γ 22,23,25, murine IFN-γ has not been produced in the plant. IFN-γ mediates its pleiotropic effects on target cells through a species-specific interaction with its receptor 13,14. As shown in figure 2, the comparison of IFN-γ amino acid sequence between mice and humans indicated that residues involved in the binding to the receptor subunit IFN-γR1 have 20% identity. Therefore, to find out the immunological response, the utilization of homologous protein seems to be necessary for experimental work on mouse model systems. In this regard, the transient plant expression system was considered a fast and effective platform for the production of recombinant mouse IFN-γ as a valuable tool for the study of IFN-γ effects in mouse models. Therefore, we investigated the relative production of bioactive murine IFN-γ along with HBsAg as a candidate vaccine in tobacco and lettuce leaves. Results of the Western blot analysis showed that the mIFN-γ was expressed on an agroinfiltrated plant and appeared to be the monomer size weighted about 25 kDa (Figure 3). The expected molecular weight of mature monomer mouse IFN-γ (without the N-terminal signal peptide) is about 15 kDa. In silico analysis showed that IFN-γ includes two potential sites for N-glycosylation in the positions 38 and 90 (Figure 2). However, in this experiment, a specific band of approximately 25 kDa was observed, suggesting that the mouse IFN-γ produced in plants underwent post-translational modifications. Although IFN-γ in non-glycosylated form is active and glycosylation is not essential for its biological activity, it is important for increasing the in vivo stability of IFN-γ 15,35. Stability enhancement is an important strategy to develop the application of recombinant IFN-γ as a potent vaccine adjuvant. This might indicate the efficiency of plant cells in mIFN-γ glycosylation during transient expression. IFN‐γ with known biological function is a noncovalent homodimer consisting of two 20-25 kDa variably glycosylated polypeptides 36. As shown in figure 3, the glycosylated molecule indicates a molecular weight of 25 kDa yielding about 60-70 kDa suggesting the probable homodimer form. Furthermore, the expression of MHC class II was actively stimulated by the plant-produced IFN-γ indicating that the recombinant mIFN-γ was correctly folded and is biologically active (Figure 5).
Additionally, we aimed to evaluate the feasibility and utility of utilizing a co-expression system for the production of these proteins. Therefore, we conducted a comparative analysis of the expression changes of mIFN-γ in various expression cassettes using ELISA. The results showed that the amount of mIFN-γ was higher in tobacco leaves agroinfiltrated with IFN-γ-TEV-HBsAg bicistronic construct, although not statistically significant (Figure 4). It is deduced that the insertion of TEV leader sequence in the upstream of the mIFN-γ gene did not increase the expression level of mIFN-γ. The result is not in agreement with our previous findings where the higher level of expression of mGM-CSF was gained in plants agroinfiltrated with monocistronic construct having TEV leader upstream of the cytokine gene 37. These indicated that in the transgene construct, in addition to the plant regulatory elements, the intrinsic characteristic of the gene of interest has impacts on the expression of transgene in the plant. The role of TEV 5ʹ leader in increasing the level of expression of monocistronic and bicistronic constructs was demonstrated in previous studies 27,38. Furthermore, the variation observed in expression level in transient transformants correlated with many parameters including the growth stage and competency of the leaves and other factors such as Agrobacterium culture and infiltration condition 39. Also as shown in figure 4, the comparative analysis of the bicistronic expression cassettes in tobacco and lettuce indicated that tobacco leaves had significantly higher mIFN-γ expression level compared to lettuce. This is consistent with other research which has shown that the transient expression levels of human growth hormone in tobacco were greater compared to lettuce leaves 40. Tobacco, as a non-food crop model plant species, has a facile and efficient transformation system and produces high leaf biomass. The limitation of using tobacco as the host for molecular farming is the presence of a variety of metabolites, including nicotine. Lettuce is another leafy crop used to produce recombinant proteins that could be directly consumed 41. Since IFN-γ given orally has biological activity, therefore plant produced IFN-γ could be used in edible vaccines.
Conclusion :
The transient plant expression system could be known as a potential and efficient production platform for vaccine research in the experimental animal phase. In the present study, the functional murine IFN-γ was successfully co-expressed in tobacco leaves along with HBsAg due to its adjuvant significance to advance the immunological response against HBsAg in preclinical studies.
Acknowledgement :
The authors greatly acknowledge Dr. Farida Nato (Pasteur Institute of Paris) for guiding the genetic molecular manipulation: Dr Geneviève Milon (Pasteur Institute, Paris, France) for supervising the cell culture research activities and Dr. Aime Nato (University of Orsay) for plant tissue culture studies.
Conflict of Interest :
The authors declare that there is no conflict of interest in this study.
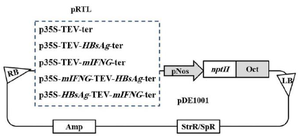
Figure 1. Schematic presentation of the gene arrangements in TEV 5ʹ leader -mediated bicistronic and monocistronic constructs that were sub-cloned from the recombinant pRTL into pDE1001 used to drive expression of mIFN-γ and HBsAg in tobacco and lettuce leaves. TEV: TEV 5ʹ leader sequence; LB and RB: left and right T-DNA borders; p35S and ter: promoter and terminator from Cauliflower Mosaic Virus; mINFG: cDNA encoding mouse-interferon gamma; pNOS and Oct: the promoter of the nopaline synthase gene and the polyadenylation signal of the Octopine Synthase (OCS) gene from the Agrobacterium tumefaciens Ti-plasmid; NPTII: kanamycin resistance gene encoding neomycin phosphotransferase II; Amp and SpR/StrR: ampicillin and spectinomycin/streptomycin resistance genes, respectively.
|
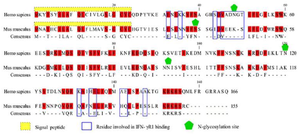
Figure 2. The amino acid sequence alignment of IFN-γ from Homo sapiens (UniProt entry P01579.1) and Mus musculus (UniProt entry P01580.1). The conserved residues are colored in red and the critical residues involved in IFN-γR1 binding are boxed in blue.
|
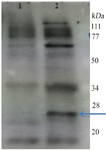
Figure 3. Western blot analysis of transient plant produced recombinant mIFN-γ using anti IFN-γ. Lane 1: control plant soluble extract, 2: transformed plant leaf extract.
|
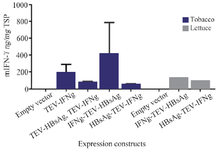
Figure 4. The comparison analysis of the expression level of mIFN-γ in tobacco and lettuce leaves agroinfiltrated with different expression cassettes by ELISA. Results are presented as means ± standard error of the mean (SEM).
|
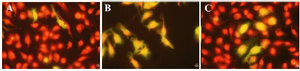
Figure 5. Immunolabeling of MHC class II molecules of macrophages from BALB/c mice bone marrow using the mAb M5/114 and anti-rat IgG F(ab′)2 coupled to FITC (green staining). Macrophages were cultured for 18 hr with (A) plant negative control, (B) recombinant mouse IFN-γ as a positive control, (C) column purified plant produced mouse IFN-γ.
|
|