Designing and Development of a Tandem Bivalent Nanobody against VEGF165
-
Khodabakhsh, Farnaz
-
Department of Genetics and Advanced Medical Technology, Medical Biotechnology Research Center, Faculty of Medicine, AJA University of Medical Sciences, Tehran, Iran
-
Salimian, Morteza
-
Department of Medical Laboratory, Kashan University of Medical Sciences, Kashan,
-
Ziaee, Pardis
-
Department of Biology, Central Tehran Branch, Islamic Azad University, Tehran, Iran
-
Kazemi-Lomedasht, Fatemeh
-
Venom and Biotherapeutics Molecules Laboratory, Department of Biotechnology, Biotechnology Research Center, Pasteur Institute of Iran, Tehran, Iran
-
 Ahangari Cohan, Reza
Department of Nanobiotechnology, New Technologies Research Group, Pasteur Institute of Iran, Tehran, Iran, Tel: +98 21 6411216, E-mail: cohan_r@pasteur.ac.ir
Ahangari Cohan, Reza
Department of Nanobiotechnology, New Technologies Research Group, Pasteur Institute of Iran, Tehran, Iran, Tel: +98 21 6411216, E-mail: cohan_r@pasteur.ac.ir
Abstract: Background: Inhibition of angiogenesis using monoclonal antibodies is an effective strategy in cancer therapy. However, they could not penetrate sufficiently into solid tumors. Antibody fragments have solved this issue. However, they suffer from short in vivo half-life. In the current study, a tandem bivalent strategy was used to enhance the pharmacokinetic parameters of an anti-VEGF165 nanobody.
Methods: Homology modeling and MD simulation were used to check the stability of protein. The cDNA was cloned into pHEN6C vector and the expression was investigated in WK6 Escherichia coli (E. coli) cells by SDS-PAGE and western blot. After purification, the size distribution of tandem bivalent nanobody was investigated by dynamic light scattering. Moreover, in vitro antiproliferative activity and pharmacokinetic study were studied in HUVECs and Balb/c mice, respectively.
Results: RMSD analysis revealed the tandem bivalent nanobody had good structural stability after 50 ns of simulation. A hinge region of llama IgG2 was used to fuse the domains. The expression was induced by 1 mM IPTG at 25°C for overnight. A 30 kDa band in 12% polyacrylamide gel and nitrocellulose paper has confirmed the expression. The protein was successfully purified using metal affinity chromatography. MTT assay revealed there is no significant difference between the antiproliferative activity of tandem bivalent nanobody and the native protein. The hydrodynamic radius and terminal half-life of tandem bivalent nanobody increased approximately 2-fold by multivalency compared to the native protein.
Conclusion: Our data revealed that the physicochemical as well as in vivo pharmacokinetic parameters of tandem bivalent nanobody was significantly improved.
Introduction :
Approximately 30% of recombinant DNA technology based drugs are antibodies that are frequently used in inflammatory diseases and cancers 1,2. Monoclonal antibodies (mAbs) could concurrently be used for di-agnostic and therapeutic purposes in medicine. However, their manufacturing is practically difficult and costly and puts an extra burden on healthcare system. In addition, because of the large size (~150 kDa), they have a poor distribution and low penetration rate into the solid tumors 3. Therefore, many efforts have been made to overcome such limitations and improve the pharmacological properties including design of antigen-binding fragments (Fab), variable fragments (Fv), single-chain variable Fragments (scFv), and single-domain antibodies (Nanobodies) 4,5. Antibody fragments have superior advantages over conventional antibodies. For example, due to the smaller size, they penetrate easily to solid tissues. Moreover, the lack of Fc domain reduces the risk of immunogenicity in humans as well as heterogeneity in the product. In the manufacturing process, they do not need mammalian expression systems which are often costly and time-consuming 6. Prokaryotic expression systems can be applied to manufacture them and usually provide much more protein than eukaryotic systems 7. Due to high solubility, low cost of production, high stability at harsh conditions, rapid penetration into solid tissues, detection of hidden antigenic sites, and lower immunogenicity with a specific affinity, the smallest binding antibody fragments (Nanobodies) have more advantages than other antibody fragments 4,8. However, they suffer from short in vivo half-life and low affinity 9. Numerous strategies were developed for enhancing pharmacokinetic parameters of small therapeutic proteins such as PEGylation and PASylation 10-14. However, each strategy has its own cons and pros. For example, PASylation technique suffers from low expression level of PASylated protein due to repeat sequences as observed in our previous study 14. Moreover, in another study, it was elucidated that the PAS sequence could decrease the affinity of PASylated protein in comparison to the native one 15. The preparation of multivalent forms of antibody fragments enhances the efficacy and in vivo distribution 16. Increasing the valency of antibody derivatives improves the pharmacokinetic properties of these fragments as well 1,17. In addition, by repeating monovalent single domain antibodies, the binding avidity could increase compared to the monomeric form 16.
Since Vascular Endothelial Growth Factor165 (VEGF165) isoform is a key factor in tumor angiogenesis and metastasis 18, therefore, in the current study, a recombinant tandem bivalent single-domain antibody against VEGF165 was designed, expressed, and purified. Finally, the size distribution and in vivo half-life of purified protein were investigated.
Materials and Methods :
3D-modeling and molecular dynamics simulation: The 3D-structure of tandem bivalent nanobody was generated using homology modeling. The crystal structure of nanobody NbFedF9 (PDB code: 4W6Y, 1.57 Å) was used as the template. One hundred models were generated by Modeller software version 9.24 19. The best model was selected based on discrete optimized protein energy (DOPE) score. The quality of the model was assessed by phi (φ) and psi (ψ) analysis 20. The stability of tandem bivalent nanobody was finally investigated by all-atom molecular dynamics simulation using GROMACS package version 2016.3 with OPLS-AA forcefield in a Linux Mint 17 operating system 21. The Extended Simple Point Charge (SPCE) water model was used for the solvent molecules. Tandem bivalent nanobody was solvated in a cubic box at a distance of 1 nm from the edges. The periodic boundary conditions were also applied. Appropriate number of Na+ and Cl- ions were added to neutralize the system. The steepest descent algorithm was used for energy minimization of the system within 200 picoseconds. The temperature and pressure of system were kept at 300°K and 1 atmosphere using Berendsen thermostat and Parrinello-Rahman barostat algorithms, respectively. The system was then subjected to the all-atom simulation until the Root Mean Square Deviation (RMSD) of the fusion protein reached to a plateau. All visualizations were performed using VMD software version 1.9.3 22.
Construct design and cloning: The anti-VEGF165 specific nanobody gene was a gift from Dr. Behdani (Pasteur Institute of Iran) and subcloned into pHEN6C vector. The nanobody gene was amplified using forward (5´-GCCCAG CCG GCC ATG GCC CAG KTG CAG CTA CAG GAG TCN GGN GG-3´) and reverse (5´-GCC TGA TTC CTG CAG CTG CAC CTG TGC CAT TGG AGC TTT GGG AGC TTT GGA GCT GGG GTC TTC GCT GTG GTG CGC TGA GGA GAC GGT GAC CTG GGT-3´) primers. The PCR program was as follows: initial denaturation at 95°C for 10 min, followed by 30 cycles of denaturation at 95°C for 40 s, annealing at 60°C for 40 s, extension at 72°C for 40 s, and final elongation at 72°C for 10 min. After the PCR purification, the product was digested with PstI and SfiI restriction enzymes and ligated into pHEN6C vector that comprises the first anti-VEGF165 nanobody 9. The accuracy of recombinant construct (VEGF165-tandem bivalent nanobody) was confirmed by sequencing.
Expression: Transformation of the recombinant plasmid to week 6 E. coli cells was performed by heat shock method. The cells were cultured on Luria–Bertani (LB) agar plates with 100 µg/ml of ampicillin and incubated overnight at 37°C. The colonies were then inoculated into 5 ml of Terrific Broth (TB) medium in a shaker incubator at 37°C until the OD600 reached to 0.5. The expression was induced by adding 1 mM of Isopropyl ß-D-1-thiogalactopyranoside (IPTG) to the culture after 16 hr. Finally, the cells were centrifuged at 4000 g for 5 min and the obtained pellets were stored at -20°C for further steps.
SDS-PAGE and western blotting: SDS–PAGE was done in a 12% (w/v) gel with a Bio-Rad instrument according to the manufacturer's protocol (Bio-Rad, USA). The protein samples were mixed with the loading buffer and boiled at 95°C for 5 min and subsequently subjected to electrophoresis. The staining of gel was done using Coomassie Brilliant Blue G-250. The gel was destained using 2.5% methanol and 10% acetic acid for 4 hr. To identify the desired protein, a western blot technique was employed using anti-His monoclonal antibody. The separated proteins were transferred onto a nitrocellulose membrane (Sigma, USA). Then, blocking step was performed using 2% skim milk in PBS. The specific detection of the desired protein was carried out by an anti-His HRP conjugated antibody (1:2000 dilution, Roche, USA). The color development for peroxidase activity was performed by DAB solution (60 mg of DAB, 100 µl of H2O2, 6 ml of methanol, and 50 ml of TBS up to 100 ml of ultra-pure water) (Sigma, USA).
Protein extraction and purification: For purification, the bacterial cultures were scaled up to 750 ml in a shaker incubator at 37°C. When the OD600 of cultures reached to 0.5, the induction was done with 1 Mm of IPTG and they were incubated for 16 hr. After harvesting the cells, the periplasmic proteins were extracted from the pellets by the osmotic method 10. The nickel-nitrilotriacetic acid (Qiagen, USA) affinity chromatography was used for purification of the periplasmic extracts. The washing step was done using 20 mM of imidazole, and then, the desired protein was eluted with 500 mM of imidazole. The purified protein was also analyzed using 12% gel polyacrylamide electrophoresis. The protein was concentrated and buffer exchanged using Amicon® ultrafilter (MWCO 3 kDa, Millipore, USA) for further characterizations. Additionally, the native protein was expressed and purified as control with the same procedures in the current study.
Size distribution measurements: Size distributions of tandem bivalent anti-VEGF165 nanobody and anti-VEGF165 nanobody were measured using Zetasizer Nano ZS (Malvern, USA) at concentration of 0.3 mg/ml.
Proliferation assay: The antiproliferative effect of nanobody and tandem bivalent nanobody was studied in vitro. HUVEC (Lonza Biologics, USA) and HEK293 (Pasteur Institute of Iran) cells were used as VEGFR2-positive 23 and -negative 24 cells, respectively. The cells were cultured in the specific media according to the instructions. Different concentrations of proteins were mixed with recombinant VEGF165 (50 ng/ml) for 4 hr and the mixture was then added to the cultured cells. After 72 hr of incubation, 50 µl of MTT solution (5 mg/ml) was added and the plates were incubated for 4 hr at 37°C. Finally, DMSO was added to the wells and the absorbance was measured at a wavelength of 570 nm using Epoch spectrophotometer (BioTeK, USA). Two-way analysis of variance (ANOVA) was used to compare the significant difference (p<0.05) between the groups using Prism 8 (GraphPad Software, Inc., La Jolla, CA).
Pharmacokinetic study: Balb/c mice (18 g, n=18) were used for pharmacokinetic study. An intravenous dose (5 mg/kg) was injected to each mouse. Blood samples were collected at different time intervals (0, 15 min, 30 min, 1 hr, 2 hr, 3 hr, 4 hr, 5 hr, and 6 hr). PBS was used as negative control. After plasma preparation, the protein concentration was measured by a home-made ELISA using anti-His monoclonal antibody as previously described 16. The terminal half-life (T1/2) and Clearance (CL) were determined using linear regression of the last three plasma concentrations.
Results :
3D-modeling and molecular dynamics simulation: Homology modeling and molecular dynamics simulation were used to check the stability of three dimensional structure of tandem bivalent nanobody. The crystal structure of nanobody NbFedF9 with a sequence identity of 72.1% was used for homology modeling. Ramachandran plot analysis (Phi and psi torsion angles) elucidated that 96.8% of residues were located in favored and allowed regions in the generated 3D-model. The RMSD analysis revealed that the 3D-model of fusion protein was stable after ~20 nanoseconds of molecular dynamics simulation in water (Figure 1).
Construct design and cloning: The nanobody gene was amplified using the primers from pHEN6-anti-VEGF165 nanobody plasmid. The PCR product was purified, digested with PstI and SfiI restriction enzymes, and ligated into pHEN6C expression vector to construct the tandem bivalent nanobody (Figure 2). DNA sequencing confirmed the accuracy of the nanobody (S1, supplementary file) and tandem bivalent nanobody (S2, supplementary file) constructs.
SDS-PAGE and western blotting: SDS-poly acrylamide gel electrophoresis showed a band at ~30 kDa in the 12% gel (Figure 3). The western blot also confirmed the expression of recombinant anti-VEGF165 tandem bivalent nanobody (Figure 4).
Protein extraction and purification: The nanobody and bivalent nanobody were extracted from the periplasmic space by the osmotic method and subjected to nickel affinity chromatography. Both proteins were successfully eluted using buffer containing 500 mM of imidazole. The bivalent protein showed a band at ~30 kDa in 12% SDS-PAGE gel (Figure 5). However, the native protein showed a band at ~15 kDa in the 12% SDS-PAGE gel (Figure 6). The proteins were finally concentrated and buffer exchanged.
Size distribution: The hydrodynamic radius of tandem bivalent anti-VEGF165 and anti-VEGF165 nanobodies was investigated by Dynamic Light Scattering (DLS). The hydrodynamic radius of tandem bivalent anti-VEGF165 nanobody increased approximately 2-fold by multivalency compared to anti-VEGF165 nanobody (Figure 7). Size distribution showed a peak at 153 nm for tandem bivalent anti-VEGF165 nanobody, while this value was 77 nm for anti-VEGF165 nanobody 14. Size distribution parameters of nanobody and tandem bivalent nanobody are summarized in table 1.
Proliferation assay: The antiproliferative activity of nanobody and tandem bivalent nanobody was studied on HUVEC and HEK293 cells. As expected, the proteins did not exhibit an anti-proliferative activity against HEK293 cells due to lack of VEGFR2 receptor on the surface cell (Data not shown). In contrast, both proteins inhibited the proliferation of HUVEC cells in a dose-dependent manner (Figure 8). Data analysis revealed that there was no significant difference between the antiproliferative activity of tandem bivalent nanobody and the native one.
Pharmacokinetic study: Pharmacokinetic study revealed that the terminal half-life (T1/2) increased approximately 2-fold after a single dose administration of the proteins (Figure 9). The measurement of pharmacokinetic parameters also elucidated that the extended in vivo half-life was obtained via a decline in the clearance of tandem bivalent nanobody (Table 2).
Discussion :
Cancer treatment using monoclonal antibodies is an ongoing practice. Monoclonal antibodies play inevitable roles in medicine and research. Most of them exert their effects by binding to transmembrane receptors or soluble ligands, interfering with signal transduction pathways, and thereby inhibiting tumor cell proliferation and angiogenesis. Because of low stability, solubility, and large size of conventional antibodies, next-generation antibodies have been developed. One approach is development of fragmented antibodies that just contain the antigen binding domain of whole antibodies. However, in some cases, such modifications remarkably affect both the function and affinity of native protein 25,26.
Development of single-domain antibodies with proper antigen-binding specificity is one of the most popular methods for fragmented antibody generation. There is a high sequence similarity between human variable heavy chains and these fragments. In addition, more hydrophilicity has occurred in these fragments due to four amino acid substitutions in framework 2 that leads to high solubility of such fragmented antibodies 27. On the other hand, their structure causes high stability and tolerability to hard conditions without losing their biological activity. Despite superior advantages over conventional antibodies, they suffer from low affinity and poor in vivo pharmacokinetics 28.
Affinity maturation and multivalency can compensate the low affinity of fragmented antibodies 29. These miniaturized antibodies also have a short in vivo half-life as these small-sized antibodies are rapidly cleared in kidney by glomerular filtration. Some investigations revealed that multivalency could concurrently improve plasma half-life and affinity of nanobodies 30. Therefore, in the current study, an anti-VEGF165 tandem bivalent nanobody was successfully designed, modeled, expressed, and purified to enhance affinity as well as pharmacokinetic parameters of the single domain antibody. The molecular dynamics simulation revealed that the designed nanobody has a good structural stability in water. The purified tandem bivalent nanobody showed a 2-fold increase in the hydrodynamic volume compared to the nanobody. Many studies demonstrated that an increase in the size of small therapeutic proteins could remarkably improve the pharmacokinetic profile via a reduction in glomerular filtration rate in kidney 31. In vitro biological activity test elucidated a dose-dependent anti-proliferative activity for nanobody and tandem bivalent nanobody against VEGFR2-expressing HUVEC cells. However, there was no significant difference between the antiproliferative activity of tandem bivalent nanobody and the native one. Moreover, the proteins did not have an anti-proliferative activity against HEK293 cells due to lack of VEGFR2 expression, as also reported in our previous studies 14,23. The designed tandem bivalent anti-VEGF165 nanobody was subjected to further in vivo characterization to evaluate the pharmacokinetic enhancement 32,33. Pharmacokinetic study indicated that the terminal half-life of tandem bivalent nanobody improved approximately 2-fold after an intravenous injection in Balb/c mice. The pharmacokinetic data also revealed that the extended terminal half-life was associated with a decrease in the clearance parameter. In a recent study, Sadeghi A et al designed a mono-specific anti-VEGF121 bivalent nanobody with extended plasma half-life for treatment of pathologic vascularization 23. In another study, Behdani et al developed an anti-VEGFR2 diabody that could potently inhibit the VEGFR2 receptor on the surface of cancer cells 34.
Conclusion :
In conclusion, recent studies indicated that multivalency can improve poor characteristics of fragmented antibodies to generate novel and potent magic tools for cancer therapy.
Acknowledgement :
The authors express their deep gratitude to all who provided support during this research.
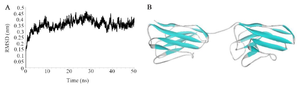
Figure 1. A) The root mean square deviations of tandem bivalent nanobody structure during 50 nanoseconds of molecular dynamics simulation. The fusion protein was reached to a plateau after ~20 nanoseconds of simulation in water. B) The 3D-structure of the designed tandem bivalent nanobody after 50 nanoseconds of molecular dynamics simulation.
|
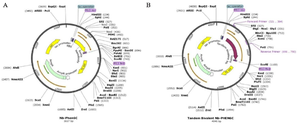
Figure 2. Schematic representation of (A) pHEN6-anti-VEGF165 nanobody and (B) pHEN6c-VEGF165-bivalent plasmids.
|

Figure 3. SDS-PAGE analysis of the expressed tandem bivalent anti-VEGF165 nanobody showed a band at ~30 kDa.
|
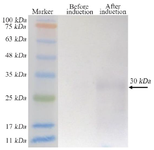
Figure 4. Western blot analysis confiremd the expression of tandem bivalent anti-VEGF165 nanobody
|
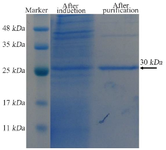
Figure 5. SDS-PAGE analysis of the expressed and purified tandem bivalent anti-VEGF165 nanobody showed a band at 30 kDa.
|
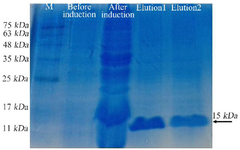
Figure 6. SDS-PAGE analysis of the expressed and purified anti-VEGF165 nanobody showed a band at 15 kDa.
|
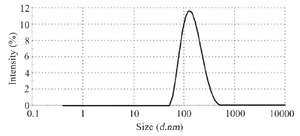
Figure 7. Hydrodynamic radius measuring tandem bivalent anti-VEGF165 nanobody using dynamic light scattering.
|
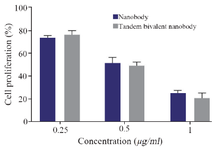
Figure 8. The antiproliferative activity of nanobody and tandem bivalent nanobody on HUVEC cells.
|
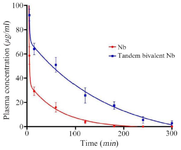
Figure 9. Plasma concentration monitoring of nanobody and tandem bivalent nanobody after injection in mice.
|

S1, supplementary file. > Nanobody
ATGAAATACCTATTGCCTACGGCAGCCGCTGGATTGTTATTACTCGCGGCCCAGCCGGCCATGGCCCAGGTGCAGCTGCAGGAGTCTGGAGGAGGCTCGGTGCAGGCTGGAGGGTCTCTGAGACTCTCCTGTGCAGCCTCTGGATTCGCCTACAGTACCTACTCCA TGGGCTGGTTCCGCCAGGTTCCAGGGAAGGAGCGTGAGGCGGTCGCAACTATCAACAGTGGTACTTTTAGGCTATGGTATACA GACTCAGTGAAGGGCCGATTCACCATCTCACGAGACAACGCCAAGAACATGCTGTATCTGCAAATGAACAGCCTGAAACCTG AGGACACGGCCATCTATTACTGTGCGGCGAGGGCCTGGTCCCCCTATAGTTCGACTGTAGACGCCGGTGACTTTCGTTACTGG GGCCAGGGGACCCAGGTCACCGTCTCCTCACACCACCATCA CCATCACTAA
|

S2, supplementary file. > Tandem bivalent nanobody
ATGAAATACCTATTGCCTACGGCAGCCGCTGGATTGTTATTACTCGCGGCCCAGCCGGCCATGGCCCAGGTGCAGCTGCAGGA GTCTGGAGGAGGCTCGGTGCAGGCTGGAGGGTCTCTGAGACTCTCCTGTGCAGCCTCTGGATTCGCCTACAGTACCTACTCCA TGGGCTGGTTCCGCCAGGTTCCAGGGAAGGAGCGTGAGGCGGTCGCAACTATCAACAGTGGTACTTTTAGGCTATGGTATACA GACTCAGTGAAGGGCCGATTCACCATCTCACGAGACAACGCCAAGAACATGCTGTATCTGCAAATGAACAGCCTGAAACCTG AGGACACGGCCATCTATTACTGTGCGGCGAGGGCCTGGTCCCCCTATAGTTCGACTGTAGACGCCGGTGACTTTCGTTACTGG GGCCAGGGGACCCAGGTCACCGTCTCCTCAGCGCACCACAGCGAAGACCCCAGCTCCAAAGCTCCCAAAGCTCCAATGGCCC AGGTGCAGCTGCAGGAGTCTGGAGGAGGCTCGGTGCAGGCTGGAGGGTCTCTGAGACTCTCCTGTGCAGCCTCTGGATTCGCC TACAGTACCTACTCCATGGGCTGGTTCCGCCAGGTTCCAGGGAAGGAGCGTGAGGCGGTCGCAACTATCAACAGTGGTACTTT TAGGCTATGGTATACAGACTCAGTGAAGGGCCGATTCACCATCTCACGAGACAACGCCAAGAACATGCTGTATCTGCAAATG AACAGCCTGAAACCTGAGGACACGGCCATCTATTACTGTGCGGCGAGGGCCTGGTCCCCCTATAGTTCGACTGTAGACGCCG GTGACTTTCGTTACTGGGGCCAGGGGACCCAGGTCACCGTC TCCTCACACCACCATCACCATCACTAA
|
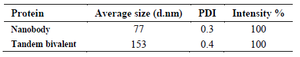
Table 1. Size distribution parameters of nanobody and tandem bivalent nanobody
|
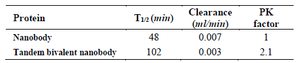
Table 2. Pharmacokinetic parameters obtained for tandem bivalent nanobody and the native one
|
|