Production of Monoclonal Antibody against Human Nestin
-
Hadavi, Reza
-
Monoclonal Antibody Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Zarnani, Amir-Hassan
-
Nanobiotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Ahmadvand, Negah
-
Monoclonal Antibody Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Mahmoudi, Ahmad Reza
-
Monoclonal Antibody Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Bayat, Ali Ahmad
-
Monoclonal Antibody Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Mahmoudian, Jafar
-
Monoclonal Antibody Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Sadeghi, Mohammad Reza
-
Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Soltanghoraee, Haleh
-
Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Akhondi, Mohammad Mehdi
-
Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Tarahomi, Majid
-
Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Jeddi-Tehrani, Mahmood
-
Monoclonal Antibody Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
 Rabbani, Hodjattallah
Ph.D., Monoclonal Antibody Research Center, ACECR, Tehran, Iran, P.O. Box: 19615-1177, Tel: +98 21 22432020; E-mail: hodrab@ki.se
Rabbani, Hodjattallah
Ph.D., Monoclonal Antibody Research Center, ACECR, Tehran, Iran, P.O. Box: 19615-1177, Tel: +98 21 22432020; E-mail: hodrab@ki.se
-
Monoclonal Antibody Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Immune and Gene Therapy Lab, CCK, Department of Oncology-Pathology, Karolinska University Hospital Solna, Karolinska Institutet , Stockholm, Sweden
Abstract: We have employed a peptide-based antibody generation protocol for producing antibody against human nestin. Using a 12-mer synthetic peptide from repetitive region of human nestin protein devoid of any N- or O-glyco-sylation sequences, we generated a mouse monoclonal antibody capable of recognizing human, mouse, bovine, and rat nestin. A wide variety of nestin proteins ranging from 140-250 kDa was detected by this antibody. This antibody is highly specific and functional in applications such as ELISA, flow cytometry, immunocytochemistry, and Western blot assays.
Introduction :
Nestin is a protein of about 240 kDa which belongs to type VI Intermediate Filament (IF) proteins. Nestin is considered as a hallmark for proliferating neural progenitor cells during early embryonic development. Although it is reported not to be expressed in adult tissues, (1) recently expression of nestin in the hair follicle stem cells has been reported (2). Nestin is evolutionary conserved within the variety of species indicating its important role in early development.
Recent advances in cancer research have revealed the expression of nestin in some cancer cells. Expression of nestin has been reported in pancreatic carcinoma (3, 4), breast cancer (5), glioblastoma (6), high-grade astrocy-toma (7), dermatofibrosarcoma protuberans (8), and also thyroid tumors (9) with different molecular weights ranging from 180-240 kDa. This variation in electerophoretic mobility is mainly because of differential post-
translational modifications like phosphorylation and glycosylation (10).
It appears that a wide variety of tumors are expressing nestin transcript without its expression at the protein level. One reason for such discrepancy might be due to the lack of a proper anti-nestin antibody recognizing all different types of nestin. To overcome this problem and also producing better research tools we decided to generate a monoclonal antibody capable of distinguishing all different variants of nestin. To do this a peptide-based antibody generation approach was selected and regions devoid of any N- or O-linked glycosylation were selected to target. These regions should also be structurally exposed to the surface of native protein for antibody binding. Regions of a protein molecule (rich in hydrophilic amino acids) tend to dissolve in water and consequently more ex-posed to outer part of molecule was selected. In this regard internet-based online databases such as Ensembl (http://www.ensembl.org/ index.html), and CBS prediction server (http: //www.cbs.dtu. dk/services/) was used.
Materials and Methods :
Peptide conjugation: The amino acid sequence of human nestin was carefully analyzed and compared to all other species. A 12-mer peptide of PEVGDEEALRPL from human nestin corresponding to amino acids 681-692 (NM_ 006617) was selected as immunogen. A cysteine residue was added to the C-terminus end of the peptide to facilitate the conjugation to carrier protein. Immunograde peptide was purchased from Thermo Electron Corpor-ation, GmbH, Ulm, Germany. Keyhole Limpet Hemocyanin (KLH)-conjugated peptide was used for generating mouse monoclonal antibody according to standard protocol with minor modifications (11, 12).
Briefly, 5 mg of Keyhole Limpet Hemo-cyanin (KLH) (Sigma, cat. no: H7017) was dissolved in 1 ml of deionized water. One mg of Maleimidobenzoyl-N-hydroxysuccinimide ester (MBS) dissolved in 200 µl of Dimethyl Formamide (DMF) was then added to the carrier protein solution. The mixture was incubated at room temperature for 2 hrs with gentle stirring followed by dialysis against large volume of Phosphate Buffer Salin (PBS) overnight. In a separate tube 5 mg of peptide was dissolved in 1 ml of Phosphate Buffered Saline (PBS). MBS-activated protein was then added to the peptide solution and the mixture was incubated for 4 hrs at room temperature. After overnight dialysis against PBS, the conjugate was stored at -20 °C for later use. All chemicals for bioconjugation were purchased from Sigma, St. Louis, USA. The same procedure was performed for conjugation of peptide to BSA.
Evaluation of conjugated peptide by SDS-PAGE: To check the efficiency of conjugation, 10 ug of peptide-BSA was mixed with 10 ul of sample buffer, boiled for 2 - 5 min and cooled on ice. Electerophoresis was performed in a 10% SDS-PAGE gel with a mini-PROTEAN electerophoresis instrument (Bio-Rad Laboratories, Hercules,CA,USA) 100 mA for 1 hr. The gel was stained with Coomassie Blue R-250 (Sigma). The change in mobility shift of conjugated BSA represented the efficiency of conjugation.
Immunization protocol: Four BALB/c mice were used for peptide immunization. Each mouse was immunized 5 times with an interval of two weeks. The first immunization was performed using Freund's complete adjuvant. Incomplete Freund's adjuvant was used for the 2nd, 3rd, 4th, and 5th immunization.
For the first immunization, 100 ug of conjugated KLH-peptide was mixed with an equal volume of Freund's complete adjuvant and injected Intra Peritoneally (IP) not exceeding 100 ul total volume. For the subsequent immunizations 50 ug of peptide-KLH were injected with Freund's incomplete adjuvant. Three days before the cell fusion, 20 ug of KLH-peptide (without any adjuvant) was injected intravenously.
Bleeding and titration of antibody: One week before the last immunization, mice were bled by a vertical incision of the tail vein. Serum ELISA assay was performed as follows: Wells of ELISA plate (Nunc, Germany) were coated with 100 ul of the immunizing peptide (20 µg/ml in PBS) at 37 °C for one hr followed by overnight incubation at 4 °C. Next day the plate was washed 3 times with PBS containing 0.05% Tween 20 (PBS-T) for 5 min. The plate was blocked with 2.5% BSA at 37 °C for 1.5 hr. Wells were then washed 3 times as above and mice sera were added to the wells in two fold serial dilutions starting from 1:100. The plate was incubated at 37 °C for 1.5 hr and washed again with PBS-T. At the next step, 100 µl of 1:1000 dilution of HRP-conjugated rabbit anti-mouse Ig (Avicenna Research Institute, Iran) was added to the wells and incubation was continued for 1.5 hr at 37 °C. After washing, 100 ul of Tetramethylbenzidine (TMB) substrate was added to each well and the plate was
incubated at room temperature in a dark place. After 15 min, the reaction was stopped by adding 30 ul of stopping solution (0.16 M H2SO4) to each well.
The Optical Density (OD) of the reactions was measured at 450 nm by an ELISA reader (BioTek, USA). Negative controls included omission of coating layer, serum (as<
Result :
Several anti-nestin monoclonal antibody producing hybridomas were obtained. Among them one clone designated as “NES-4G10G8” had a very high reactivity with immunogenic peptide in ELISA assay. Further characterization of this antibody showed that it is of an IgG1 isotype with a kappa light chain. Due to stronger reactivity of this clone in ELISA assay, all subsequent tests were carried out with this clone.
Cell lines of human, mouse, and bovine were tested by western blot (Figures 1A, B and C), immunofluorescent staining (Figures 2A, B and C) RT-PCR (Figure 3) and flow cytometry (Figure 4) for expression of nestin at both gene and protein levels. Results of RT-PCR showed that except CHO, T47D and PC3, all cell lines express nestin-specific message. Based on the sequences amplified, band sizes differed accordingly.
In western blot analysis, nestin-specific bands ranging from 140-250 kDa were observed (Figure 1). More interestingly, the pattern of bands in reducing and non-reducing conditions differed almost in all cell lines examined. This was especially the case for 140 kDa band which appeared or became stronger under reducing condition. To examine the expression pattern of nestin, cell lines were subjected to immunofluorescent staining. The results clearly showed that our antibody strongly reacts with nestin intracellulary (Figures 2A, B, C and 4).
In line with RT-PCR and western blot analysis, PC3, T47D and CHO cell lines failed to express nestin, while other cell lines showed excellent reactivity with nestin-specific monoclonal antibody.
In some settings, anti-nestin antibody was conjugated with Alexa fluor 568 and used for direct staining of cell lines. As shown in figure 2B, bovine sertoli cells and A172 cell line (as positive controls) strongly expressed nestin, while CHO cell line showed no reactivity. Summary of nestin expression in different cell lines employing three readout systems are depicted in table 2.
To further characterize our antibody, bovine sertoli cells were subjected to intracellular staining and tested by flow cytometry. The results showed that 73% of the cells were positive for expression of nestin (Figure 4).
Discussion :
Peptide-based antibody generation gave rise to identification of several hybridomas producing anti-nestin antibody. One antibody of IgG1 isotype was generated which showed to be very specific to nestin. The antibody could react with nestin protein in both non-reducing and reducing conditions with some variations. Examples of such variations are prostate cancer cell line PC3 and melanoma cell line ESTBAD-75 which did not react in non-reducing conditions but could react in reducing conditions as well as in ICC experi-ments (Figures 1A and 2A).
This variation might be due to recognition of antigenic epitope in different conformation by the antibody. Apparently this antibody recognizes a linear epitope which is structurally folded and appears and exposes to the antibody upon dissociation by 2-mercaptoethanol (reducing conditions). Furthermore, reactivity of this antibody to nestin from pancreatic carcinoma cell line PaCa2 at both reducing and non-reducing conditions (Figure 1A), indicates that the structure of nestin expressed by PaCa2 cells might be different with the structure of nestin expressed by A172 or ESTBAD-75 cell lines. This discrepancy might be due to altered Post-Translational Modification (PTM) main-ly glycosylation of nestin in cancer cells, (13) which affects the electerophoretic mobility of protein. Our results showed expression of nestin with different molecular weight ranging from 140-250 kDa confirming the previously described nestin variants (10).
Our results are in accordance to the results on expression of nestin in the literature. The reference data are shown in table 1. The only case which differs from the literature is expression of nestin in prostate cancer cell line PC3. Unlike reporting expression of nestin at only gene level in PC3 cell line (14), we did not detect expression of nestin in this cell line (in contrast to our data).
The NES-4G10G8 antibody detects nestin from other species. This might imply recogni-tion of a common epitope. Amino acid sequence alignment of the immunogenic peptide of human, bovine, mouse, hamster, pig, horse, dog and rat showed that the consensus sequence of XEXEXQEXXRPL in which glutamic acid E (2), arginine R (10), and leucine L (11) might represent the potential epitope recognized by anti-nestin clone 4G10G8 (Figure 5).
If this is true, then any nestin protein from other species containing such epitope can potentially be recognized by this antibody. So, hypothetically the horse, dog, and rat´s nestin might be recognized by this antibody which needs further investigations. This anti-body could be used as a powerful tool in stem cell studies as it is reacting in a wide variety of applications like western blot, ELISA, flow cytometry, and immunocytochemistry.
Our results show that this antibody is capable of recognizing nestin in both normal and pathological conditions as well as with possible structural diversities.
Acknowledgement :
This work was supported by grants from Ministry of Health and Medical Education of the Islamic Republic of Iran. We thank Dr. Mazdak Salavati, Ebrahim Torkabadi, and Afshin Namdar for their technical assistance.
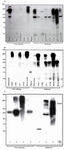
Figure 1. Western blot assay using 3 ug affinity-purified anti-nestin (4G10G8). A) Cell lysates from a human glioblastoma multiforme (GBM) and several human malignant cell lines were run in a 6% SDS-PAGE under reducing and non-reducing conditions. B) Cell lysates and several cell lines from other species were also analyzed. Several bands ranging from 150-300 kDa was observed. C) Cell lysates from 1- Rat hippocampus, 2- Mouse brain (12 days embryo), 3- Bovine brain (24 days embryo), 4- Human cerebral tumor
|
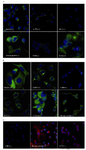
Figure 2. Immunocytochemistry (ICC) assay using 5 ug of affinity-purified anti-nestin (4G10G8). FITC-conjugated sheep anti-mouse antibody was used as secondary antibody. DAPI was used for staining the nucleus (blue). The green fluorescence represents nestin expressed in cytoplasm and the blue color re-presents the nucleus. A) Human cancer cell lines. B) Cell lines derived from other species and also a primary sertoli cells. C) To confirm the ICC results, a negative (CHO), a strong positive (bovine sertoli cells), and a weak positive nestin expressing cells were stained with anti-nestin 4G10G8 conjugated with Alexa Fluor 568 (Invitrogen). Red immunofluorescence re-presents nestin
|
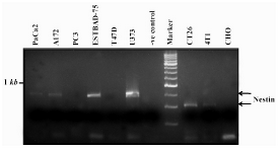
Figure 3. Expression of nestin in different cell lines by RT-PCR. The standard marker is DNA molecular weight marker XVI from Roche. The negative (-ve) control was PCR reaction without any template
|
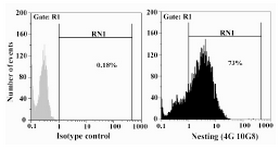
Figure 4. Indirect intracellular staining of bovine sertoli cells and flow cytometry analysis using anti-nestin (4G10G8) antibody. FITC-conjugated rabbit anti-mouse was used as secondary antibody. An irrelevant mouse IgG1 (ACECR, Tehran, Iran) was used as isotype control
|

Figure 5. The sequence of human nestin immunogenic peptide was aligned with corresponding regions of nestin protein from other species. The conserved amino acids are highlighted in gray and potential antigenic epitope is underlined
|
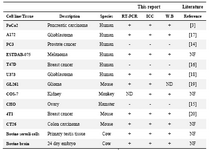
Table 1. Summary of expression profile of nestin in different cell lines
ICC=Immunocytochemistry, W.B=Western blot, ND=Not determined, NF=Not found in the literature
|

Table 2. Nestin species-specific primers
F=Forward, R=Reverse
|
|