Preliminary Study on Cost-effective L-Tryptophan Production from Indole and L-Serine by E. coli Cells
-
Sadeghiyan-Rizi, Tahereh
-
Department of Pharmaceutical Biotechnology, Faculty of Pharmacy, Isfahan University of Medical Sciences, Isfahan, Iran
-
 Fooladi, Jamshid
National Laboratory of Industrial Microbiology, Department of Biology, Faculty of Science, Alzahra University, Tehran, Iran, Tel: +98 21 22142964, E-mail: jfooladi@alzahra.ac.ir
Fooladi, Jamshid
National Laboratory of Industrial Microbiology, Department of Biology, Faculty of Science, Alzahra University, Tehran, Iran, Tel: +98 21 22142964, E-mail: jfooladi@alzahra.ac.ir
-
National Laboratory of Industrial Microbiology, Department of Biology, Faculty of Science, Alzahra University, Tehran, Iran
-
Sadrai, Sima
-
Department of Pharmaceutics, Faculty of Pharmacy, Tehran University of Medical Sciences, Tehran, Iran
Abstract: Background: L-tryptophan is used widespread in the pharmaceutical industry. The majority of L-Trp production depends on microbial processes that produce L-tryptophan from indole and L-serine. These processes are very costly due to the costs of precursors, especially L-serine. Use of inexpensive substitutions as the L-serine source of L-tryptophan production enables us to reach a cost-effective process. In this paper, effect of Triton X-100 on L-Trp production and the ability to use Iranian cane molasses as inexpensive L-serine source was investigated.
Methods: Escherichia coli (E. coli) ATCC 11303 cells were grown in 10-L fermenter containing minimal medium supplemented with beet molasses as an inexpensive carbon source and indole as tryptophan synthase inducer. Whole cells of stationary phase were used as biocatalyst for L-Trp production. Triton X-100 addition to the production medium as indole reservoir was investigated. Then, cane molasses was used as L-Ser source in L-Trp production medium. Amount of L-Tryptophan and theoretical yield of L-Trp production was determined by HPLC and by a colorimetrically method on the basis of remaining indole assay, respectively.
Results: As a result, triton X-100 increased L-Trp production three times. Also, the result showed that 0.68 mM L-Tryptophan was produced in the presence of cane molasses at 37oC for 8 hr.
Conclusion: This result showed that cane molasses of Qazvin sugar factory includes significant amounts of L-Ser that makes it a suitable substitution for L-Ser in L-Trp production. Therefore, it has the potential to be used for cost-effective L-Trp production in industrial scale.
Introduction :
L-Trp is an important ingredient in medicines and animal feeds as an essential amino acid 1-3. Its use in infusion solution form as an antidepressant, sedative and in treatment of schizophrenia and alcoholism are some of the more important applications 4,5.
Several microbial processes used for L-Trp production include direct fermentation from carbohydrates or hydrocarbons 6,7, enzymatic reaction from L-Trp precursors 8 and bioconversion from L-Trp precursors 9. Many commercial processes employ Escherichia coli (E. coli) cells to convert indole and L-Ser to L-Trp by tryptophan synthase (TSase) 10. This process is very simple and a one-step reaction catalyzed by tryptophan synthase and when precursors are employed, the complete biosynthetic pathway of L-Trp isn’t necessary and the com plicated mutations of microorganisms which control the regulatory mechanism are often not required 2. One disadvantage of this process is that precursors especially L-Ser and Pyridoxal Phosphate (PLP) are expensive. TSase is a PLP dependent enzyme. As established in our previous study, exogenous PLP isn’t needed in our reaction conditions because beet molasses contains relatively large amounts of sucrose, amino acids and vitamins such as pyridoxine and when used as the carbon source of culture medium not only accelerated the bacterial cells growth but also these cells gained a significant amount of PLP from the culture medium and when used as a biocatalyst in the production medium, they didn’t require exogenous PLP as a cofactor of TSase unlike previous reports 11. This finding is important from an economical point of view. Therefore, in the present study, the amount of intracellular PLP was investigated. Another substrate, L-Ser, is very expensive because a racemic mixture is formed during its manufacture.
To solve this problem, new methods in both process design and cheap precursor substitution have been developed 12,13. Another disadvantage of this process is high concentration of indole inhibits TSase activity. To address this matter, organic solvents were used as reservoirs of indole in production medium 2.
The aim of this research was cost-effective production of L-Trp. Therefore, E. coli ATCC 11303 was cultured in the inexpensive medium and bacterial cells induced for TSase activity during growth by addition of indole to the culture medium harvested from growth medium were transferred to production medium as biocatalyst of L-Trp production from indole and L-Ser. The possibility of Triton X-100 use as an indole reservoir to prevent inhibition of TSase by indole and Qazvin cane molasses as a cheap L-Ser substitution for cost-effective L-Trp production was investigated.
Materials and Methods :
Chemicals: Indole, L-serine, PLP, L-tryptophan, Triton X-100, TLC plates (60F254) and methanol were purchased from Merck (German), and all other chemicals were of analytical grade or the highest purity commercially available. Cane and beet molasses were obtained from Iranian sugar industries and cane and beet molasses composition is presented in table 1. Orumiyeh beet molasses was sterilized separately at 121°C for 15 min for preparation of culture medium and Qazvin cane molasses was passed through a 0.22 μ Millipore filter for preparation of reaction mixture.
Microorganism and culture conditions: E. coli ATCC 11303 was used in this research. For large scale production of biomass, a 500 ml culture, grown in complete medium containing glucose, 20 g/l; yeast extract, 25 g/l; (NH4)2SO4, 0.5 g/l in 2 L Erlenmeyer flask on a rotary shaker, 180 rpm at 37oC for 10 hr, was inoculated into 10 L of a medium containing K2HPO4, 7 g/l; KH2PO4, 3 g/l; Na3. C6H8O7.3H2O, 0.5 g/l; MgSO4.7H2O, 0.1 g/l; Orumiyeh (northwest of Iran) beet molasses, 16.2 g/l and indole, 0.058 g/l. The bioreactor was a 10 L fermenter (type MS-F1-S-10l) equipped with a draft tube and driven by a specially constructed propeller at 250 rpm and the pH of culture medium was adjusted automatically to 7.0±0.2 by titration with NaOH 10 N and HCl 1 N.
Determination of cell growth: The growth of microorganism was determined periodically by measuring the optical density at 620 nm every one hour and also 1 ml of culture medium was centrifuged at 8000 rpm at 4oC for 20 min and the harvested cell mass was dried at 105oC for 24 hr to a constant weight 14,15.
Tryptophan production: Biomass harvested from culture medium (3 g) was transferred to 500 ml Erlenmeyer flask containing production medium: 100 ml potassium phosphate buffer (0.1 M, pH=8), indole (0.05 g) and L-Ser (0.05 g), incubated on rotary shaker (180 rpm) at 37oC. After 4 hr of incubation, the production medium was centrifuged at 12000 rpm for 20 min in 4oC and the supernatant fluid was used for assay of produced L-Trp and remaining indole.
L-Trp was determined by HPLC in a Waters system equipped with isocratic pump and ultraviolet detector, by using an RP-18 column (MZ-analytical column, 4×100 mm) with 3 µm particle size and the mobile phase was phosphate buffer (0.05 M, pH=4.2): methanol (70:30). Injection volume, flow rate and λ absorbance were 50 µl, 1.2 ml/min and 220 nm, respectively 16,17. For sample analysis, L-Trp was dissolved to a concentration of 1 mg/ml in purified water as stock solution. L-Trp concentrations of 5, 4, 2, 1, 0.5, 0.2, 0.1 and 0.05 µg/ml were prepared in purified water from stock solution and used for standard curve drawing. L-Trp concentration in the supernatant fluid of production medium was determined according to standard curve. AT 3000 Autochor software was used for HPLC data analysis.
Consumed indol and the theoretical yield of L-Trp production was determined colorimetrically on the basis of remaining indole in the supernatant fluid of production medium at 490 nm by spectrophotometer 18. Briefly, indole concentrations of 0.1-1 μmol/ml were prepared with 96% ethanol and 0.5 ml of each concentration was transferred to the screw cap tube separately and 0.5 ml NaNO2 and 2 ml Gly-HCl buffer (pH=2.6, 0.1 M) were added to it. All tubes were put to the beaker containing boiling water for 5 min. Following cooling down, the absorbance of them was measured at 490 nm and used for the standard curve. Indole concentration in the supernatant fluid of the production medium was determined according to the standard curve.
Determination of intracellular PLP: Bacterial cells (3 g) were sonicated at 100% amplitude for 6 min and centrifuged at 12000 rpm at 4oC for 20 min, and then the supernatant fluid was used for determination of the intracellular PLP by TLC. PLP concentration of 0.0001 g/ml was prepared in purified water as standard solution and kept at 4oC. TLC was performed on silica gel 60F254 plates with dioxane: deionized water 1:1 (v/v) as mobile phase. Before use, the plates were activated at 50oC for 30 min in an electrically controlled oven. After activation, the plates were cooled at room temperature and stored in a closed chamber at room temperature. Samples (30 µl) were applied to the plate by use of Hamilton syringe, 1 cm above the lower edge of the plate. The spots were left to dry completely and the plate was then developed by the one-dimensional ascending technique in a rectangular chamber pre-equilibrated with the mobile phase for 15 min. After development, the plate was dried at 50oC in an oven and the spots of PLP were visualized in 254 nm UV light 19,20.
Investigation of triton X-100 effect on L-Trp production: To avoid inhibition of TSase by indole, Triton X-100, nonionic detergent, was used in the production medium as reservoir of indole. This effect was investigated by use of the 3 production media, one of them without Triton X-100 (control sample), and 2 another production media with 300 µl Triton X-100 and in one of them Triton X-100 was added at first (sample 1) and in another, it was added in 2 hr after production medium incubation (sample 2).
Investigation of L-Serine content of Qazvin cane molasses: L-Ser in molasses has been investigated by TLC and TLC scanning. 5 mg Qazvin cane molasses was diluted in 10 ml distilled water, mixed with n-butanol (1:1), vortexed, incubated at room temperature so that two phases were formed then the organic phase (upper layer) was harvested. Following evaporation, the residue was mixed with 10 ml n-butanol and 50 μl of it was loaded on the silica gel 60 F254 plate. The chromatogram was developed in rectangular glass chamber containing mobile phase n-butanol/acetic acid/water (65:13:22 V/V/V). The chromatogram was dried at 60°C for 10 min and the ninhydrin reagent solution was sprayed on the TLC and dried at 110°C for 5 min. The identification was confirmed by Rf obtained by the comparison of L-Ser standard. Then TLC scanning was performed in the wavelength of 254 nm.
Investigation of L-Trp production by Qazvin cane molasses: Qazvin cane molasses (center of Iran) capability to be used as L-Ser source of production medium was investigated. L-Ser of production medium was displaced with Qazvin cane molasses (5 g/100 ml of production medium). Because other components except L-Ser might exist in processed molasses and have an effect, reaction production medium was incubated for longer time i.e. 10 hr and sampling was done at 4, 8 and 10 hr. Since cane molasses might contain L-Trp and interfere with the results, L-Trp existing in production medium was calculated before production medium incubation (T0).
Statistical method: The experiments were carried out in triplicate. All the results were analyzed statistically by one-way analysis of variance and Tukey test with 95% confidence level using SPSS 20.0.
Results :
Determination of cell growth: According to the results, E. coli cells reached to the exponential phase of growth after 6 hr of incubation in the fermenter and introduced to the stationary phase (Figure 1). The bacterial biomass was recovered and used as a biocatalyst in the L-Trp production medium. The wet weight of harvested biomass was 105 g.
Investigation of triton X-100 effect on L-Trp production: As shown in figure 2, Triton X-100 is a suitable indole reservoir and prevents inhibition of TSase by indole. L-Trp production is increased 3 times by use of Triton X-100.
Determination of intracellular PLP: As shown in the previous study, no exogenous PLP was required as a cofactor of TSase due to the cellular uptake of PLP from the molasses during culture in the fermenter 11. This probability was investigated and results showed that there was adequate amount of PLP in the cells (Figure 3).
Investigation of L-Serine content of Qazvin cane molasses: Results obtained from scan TLC established that there was about 0.025 g L-Ser in the 5 g of Qazvin cane molasses (Figure 4).
Investigation of cane molasses usage instead of L-Ser: It was indicated that cane molasses of Qazvin sugar factory includes significant amounts of L-Ser and 0.68 mM L-Trp was produced from it as the L-Ser source of reaction (Table 2).
Discussion :
As described above, L-Trp production from indole and L-Ser was carried out using whole cells of E. coli ATCC 11303. The cost-effective L-Trp production has gradually attracted the attention of the amino acid industry 10. The following conditions are very critical for cost-effective L-Trp production via biotransformation: 1) inexpensive medium for microorganism cultivation, 2) inexpensive substitutions for precursors, 3) a short reaction time 2.
In this research, beet molasses was used as an inexpensive carbon source of culture medium. Beet molasses contains relatively large amounts of sucrose, amino acids and vitamins such as pyridoxine 20. On this reason, in our study, use of beet molasses as carbon source of culture medium not only decreased the cost of microorganism cultivation but also bacterial cells gained significant amount of PLP from culture medium, as shown in figure 3, and when used as a biocatalyst in the production medium, exogenous PLP as cofactor of TSase 11 was not required whereas in previous reports exogenous PLP had been needed 1,2,7,10, 21-25 and this finding is very important from the economic point of view.
The inhibition of TSase by indole is a major problem, so nonaqueous organic solvents or nonionic detergents are used as indole reservoirs. Results showed that L-Trp production is increased by use of Triton X-100 (Figure 2), presumably due to the increase in bacterial cells permeability and the gradual supply of indole to the reaction mixture of L-Trp production.
As the main result, L-Trp (0.68 mM) was produced from Qazvin cane molasses as L-Ser source unlike Fariman and Karaj cane molasses in our previous study 11. Cane molasses composition highly varies and depends on cane varieties, climate and processes of sugar production 26. However, it was observed that the use of cane molasses as L-Ser source of production medium showed a 32% decrease in L-Trp production in comparison to the time when L-Ser was used (Table 2). The result was expected beforehand because beet molasses contained considerable amounts of L-Trp (0.018 mM, Table 2) and L-Trp is a strong inhibitor of L-Trp production. In the report presented by Mateus (1996), an initial L-Trp led to decrease in its initial production rate 23.
Conclusion :
In this research it was proved that L-Trp production is increased by the use of Triton X-100 as indole reservoir in the production medium and also it was established Qazvin cane molasses includes significant amounts of L-Ser that makes it a suitable substitution for L-Ser in L-Trp production. This is the preliminary study on cost-effective L-Trp production by Iranian cane molasses and these results can be used in the industrial scale following further researches.
Acknowledgement :
We gratefully acknowledge the support of the National Laboratory of Industrial Microbiology at Alzahra University and Biopharmacy Laboratory of Tehran University of Medical Sciences.
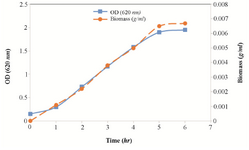
Figure 1. Growth curve ( ) and biomass curve ( ) of E. coli cells growing in the culture medium in the fermenter.
|
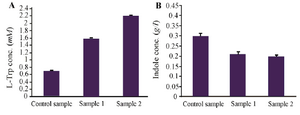
Figure 2. Effect of Triton X-100 on L-Trp production (a) and indole consumption (b).
|
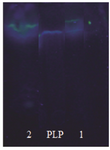
Figure 3. Determination of intracellular PLP. TLC patterns of standard sample (PLP), intracellular PLP (1), and PLP in beet molasses (2).
|

Figure 4. Investigation of L-Serine content of Qazvin cane molasses. TLC patterns of L-Ser in the Qazvin cane molasses (QCM), L-Trp standard sample (L-Trp) and L-Ser standard sample (L-Ser).
|
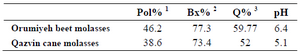
Table 1. Iranian cane and beet molasses composition
Sucrose content (g/100 ml molasses)
Total soluble solids (w/w)
Purity percentage: (Pol/Bx) × 100
|

Table 2. L-Trp production in the presence of processed molasses as L-Ser source
* Existing L-Trp in Qazvin cane molasses was measured in the t0.
|
|