Cloning and Optimization of Soluble Vascular Endothelial Growth Factor165 Expression in Escherichia coli
-
Salimi, Ali
-
Monoclonal Antibody Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
 Babashamsi, Mohammad
Monoclonal Antibody Research Center, Avicenna Research Institute, ACECR, Tehran, Iran, Tel: +98 21 22432020, E-mail: babashams@avicenna.ac.ir
Babashamsi, Mohammad
Monoclonal Antibody Research Center, Avicenna Research Institute, ACECR, Tehran, Iran, Tel: +98 21 22432020, E-mail: babashams@avicenna.ac.ir
-
Monoclonal Antibody Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
Abstract: Background: Vascular Endothelial Growth Factor (VEGF) is a coordinate regulator of physiological angiogenesis during embryogenesis, skeletal growth and reproductive functions. There are several types of VEGF, including VEGF165. VEGFs stimulate endothelial cell growth, angiogenesis, and capillary permeability. Low induction temperature is a major factor for expression of the recombinant VEGF165 in soluble form. The purpose of this study was cloning and optimization of soluble vascular endothelial growth factor165 expression in Escherichia coli (E. coli).
Methods: In this study, total RNA of HeLa cell [cervix epithelium] was extracted. The VEGF165 gene was amplified by reverse transcription polymerase chain reaction (RT-PCR), and then VEGF165 was subcloned into prokaryotic expression vectors pET-32a(+) and transformed into BL21 (DE3) E. coli strain. VEGF165 expression was optimized by fine adjustments such as induction time and incubation temperature. VEGF165 was analyzed by DNA sequencing prior to expression and the protein was further characterized by SDS-PAGE and immunoblotting using His•tag specific polyclonal antibody.
Results: Our results demonstrated that VEGF165 was successfully cloned and expressed in pET-32a(+) vector. Optimization of the expression procedure showed that, induction by 1 mM IPTG at OD600=0.7 and overnight incubation at 22oC resulted in the highest expression levels of soluble VEGF165.
Conclusion: In this study, the expression of VEGF165 in a high soluble level was successfully cloned and optimized.
Introduction :
Angiogenesis occurs in an environment where angiogenic signaling overcomes antiangiogenic factors that regulate blood development within an organ 1. Vascular Endothelial Growth Factor (VEGF) is a signal protein produced by cells that stimulates vasculogenesis and angiogenesis 2. It is part of the system that restores the oxygen supply to tissues when blood circulation is inadequate 3. Serum concentration of VEGF is high in bronchial asthma and diabetes mellitus and generally VEGF over-expression is observed in various diseases 4. VEGF's normal function is to create new blood vessels during embryonic development, new blood vessels after injury, muscle following exercise, reproductive functions and new vessels (collateral circulation) to bypass blocked vessels 5. There are multiple isoforms of VEGF-A that result from alternative splicing of mRNA from a single, 8-exon VEGF-A gene 6. These are classified into two groups which are referred to according to their terminal exon (exon 8) splice site: the proximal splice site (de-noted VEGFxxx) or distal splice site (VEGFxxxb).
In addition, alternate splicing of exon 6 and 7 alters their heparin-binding affinity and amino acid number (in humans: VEGF121, VEGF121b, VEGF145, VEGF165, VEGF165b, VEGF189, VEGF206 7. VEGF165 stimulates endothelial cell growth, angiogenisis, and capillary permeability. VEGF165 has also been twisted in pathological angiogenesis joined with tumors 8. The properties of native VEGF closely correspond to those of VEGF165. Expression of recombinant proteins in Escherichia coli (E. coli) is severely influenced under various conditions. In general, conditions that decrease the rate of protein synthesis, such as low induction temperatures tend to increase the percentage of target proteins found in soluble form 9,10. In many applications, it is desirable to express target proteins in their soluble, active form 11. The purpose of this study was cloning and optimization of soluble VEGF165 expression in E. coli.
Materials and Methods :
Materials: Reagents and materials were purchased from the following sources. HeLa cell line (Cell Bank of ARC, Iran), Easy-spin RNA Extraction kit (INtRON, USA), cDNA Reverse Transcription Kits (Novagen, USA), gel extraction kit (Qiagen, USA), the E. coli host strain BL21 (DE3), pET-32a(+) plasmid (Novagen, USA), PCR reagents, restriction enzymes and T4 DNA ligase (Promega, USA), ECL Supersignal system (Pierce, USA), plasmid extraction kit and PCR purification kits (Bioneer,USA), Trypsin-EDTA, RPMI 1640 medium and Fetal Bovine Serum (Gibco, USA), anti-His6-Peroxidase (Roche, Germany), VEGF165 Protein (Novus Biologicals, USA), PVDF Western Blotting Membranes and Skim milk (Roche, Germany). Other chemicals were all commercially available reagents of analytical grade. All solutions were prepared using deionized water.
Bioinformatic tools for PCR primer design: The PCR primers were designed by Gene Runner 3.05 Version, NEB Cutter, Codon Usage calculator and Mutation Surveyor V3.30 (Demo). Primers were designed by addition of 2 nucleotides for BamHI and 3 nucleotides for XhoI on either side of their recognition site.
Cell culture: HeLa cells were cultured in RPMI 1640 containing 10% heat inactivated fetal bovine serum, 100 units/ml penicillin G sodium, 100 µg/ml streptomycin sulfate in a 5% CO2 atmosphere at 37oC. HeLa cells were detached with 1 ml of 0.25% Trypsin-EDTA for 5 min. A density of 1×105 cells was used for RNA extraction.
RNA extraction and cDNA synthesis: Total RNA was extracted using the Easy-spin RNA Extraction kit. RNA purity was estimated by measuring the ratio of absorbance at 260 per 280 nm (approximately 1.8) and by Ethidium bromide staining of 18S and 28S RNA after agarose gel electrophoresis. Subsequently, RNA concentrations were determined using A260 values. cDNA synthesis was performed using high Capacity cDNA Reverse Transcription Kits. Two micrograms of RNA were reverse transcribed using 10 µl of 10X RT buffer, 0.8 µl of 25X dNTP Mix (100 mM), 2 µl of RT Random Primers, 1 µl of MultiScribe™ Reverse Transcriptase, and 4.2 µl of Nuclease-free H2O. Reverse transcription reaction was conducted at 37oC for 120 min. Thereafter, the reaction mixture was heated to terminate the reaction at 85°C for 5 min. Beta actin was used as a loading control in PCR for confirming cDNA. The cDNA was stored at -20°C until analysis.
Construction of VEGF165 expression vector: The VEGF165 cDNA sequence was amplified by PCR using the primers 5'-CGGGATCCCGTATGAA CTTTCTGCTGTCTTGGGTG-3' and 5'-CCCTCGAG GGTCACCGCCTTGGCTTGTCAC-3'. PCR was performed in a 25 µl master mix containing PCR 10X buffer, 1.5 mM MgCl2, 0.4 mM dNTPs, 1 unit of DNA polymerase, and 200 nmol of each forward and reverse primer. The thermal cycling parameters used for PCR were as follows: 30 s at 61oC for annealing, 30 s at 72oC for extension, and 30 s at 94oC for denaturation. After 40 cycles, amplified cDNA products were electrophoresed on a 1.5% Low Electroendosmosis (LE) agarose gel, extracted and purified by QIAquick gel extraction kit according to the manufacturer’s protocol. The gel extract was digested using BamHI and XhoI and ligated into the multiple cloning sites of pET-32a(+). The engineered VEGF165_pET-32a(+) vector was transformed into DH5α and BL21( DE3) cells. The BL21 (DE3) was cultured on LB agar plate and a colony was selected for colony PCR confirmation test.
Expression of VEGF165: Escherichia coli (E. coli) BL21 (DE3) harboring the VEGF165_pET-32a(+)positive colony was grown in 100 ml LB broth at 37°C including 100 μg/ml ampicillin to achieve an optical density (OD) of 0.7 (stationary growth phase) at 600 nm and at this time, 1 mM isopropyl-ß-D-thiogalactoside (IPTG) was added to 50 ml LB broth and the same amount was considered as uninduced (-IPTG) control. Incubation was continued at 37oC. The samples from induced and uninduced culture were collected within 2, 4, 6 hr and overnight after induction. Samples were accumulated, centrifuged at 6000 rpm for 5 min. Harvested cells were re-suspended with 6x loading protein buffer containing 2-ME, boiled and subjected to SDS-PAGE. The expression of recombinant VEGF165 protein in bacteria was confirmed by immunoblotting technique. The rate of VEGF165 expression was determined by AlphaEase FC Software. In this method, the gel was scanned and the ratio of the VEGF165 band intensity to the total band intensity was calculated (Table 1).
Western blot analysis: Whole cell pellets were separated on 12% SDS-PAGE gel and transferred to PVDF. Membranes were blocked for overnight with 5% skim milk in 0.1% Tween 20 in PBS buffer, incubated at room temperature for 2 hr with monoclonal anti His6 conjugated with horseradish peroxidase, then washed for 10 min three times and developed by supersignal chemilumines-cence substrate.
Optimization of VEGF165 expression: To look for culture parameters affecting the expression and aggregation protection of the fusion protein including time and temperature, 50 ml of the cultured BL21 (DE3) pellet under different induction times (4 and 18 hr), and temperatures (22 and 37oC) was re-suspended by swirling and sonication on ice using a microtip with the power level set of 4, at 40% duty for 20 bursts. The lysate was centrifuged for 10 min at 14,000×g to separate the soluble and insoluble fractions. The soluble and insoluble fractions were immediately mixed by 6x protein loading buffer containing 2-ME and heated for 3 min at 85oC to denature proteins. The fractions were analyzed by SDS-PAGE to look for optimal condition for VEGF165 expression. The rate of soluble VEGF165 expression was determined by AlphaEase FC Software. The gel was scanned and the ratio of the soluble VEGF165 band intensity to the insoluble VEGF165 band intensity was calculated (Table 1).
Results :
RNA extraction and cDNA synthesis: The 18 S and 28 S bands were identified from RNA extract of HeLa cells on agarose gel. Two bands were identified from PCR product of cDNA on agarose gel (Figure 1A). The band (441 bp) shorter in length was confirmed as VEGF121 while the longer band (576 bp) was normal VEGF165 as examined by sequencing.
Construction of VEGF165 expression vector: The VEGF_165pET-32a(+) was constructed with a fusion partner of Trx. The cloned sequencing showed the insertion of the VEGF165 gene into the vector (Figure 2). The results of the effect of the colony PCR test on the VEGF_165pET-32a(+) construct is shown in figure 1B. The 576 bp band confirms the perfect cloning of VEGF165. The result of obtained DNA sequence was subjected to BLAST analysis as shown in figure 3. BLAST analysis did find 100% homologous sequences in GeneBank database.
Expression of VEGF165: The protein expression was carried out under induction and un-induction conditions in different times (2, 4, 6, hr and overnight) in LB medium in shake flask. The expressed protein was subjected to 12% (w/v) SDS-PAGE under reducing conditions and visualized by Coomassie blue staining. As shown in figure 4A, there is an over expression (40.2% of the total expressed protein) after 4 hr of induction (Table 1).
Western blot analysis: The expressed VEGF165_pET-32a (+) was analyzed by Western blot method (Figure 5). The result indicates that the molecular size of VEGF165 with tags was 38 kDa (as estimated in 12% SDS–PAGE gel).
Optimization of VEGF165 expression: The effects of induction parameters (Time and temperature) on soluble VEGF165 expression were examined after addition of 1 mM of IPTG. The results indicated that the soluble VEGF165 expression was highest at 22oC. In addition, the optimal induction was performed at 18 hrs after addition of the IPTG. As shown in figure 4B, induction for 18 hr increased the soluble VEGF165 expression to 20.8% of the total expressed VEGF165 protein compared with that of 4 hr at 37oC (12.8% of the total expressed VEGF165 protein) (Table 1). Taken together, the optimal culture condition for VEGF165 expression by E. coli was established as 1 mM of IPTG at 22oC for 18 hr.
Discussion :
VEGF-A has several variants; VEGF165 appears to be the most abundant and potent isoform, followed by VEGF121 and VEGF189 12,13. Isoforms other than VEGF121 contain basic heparin-binding regions and are not freely diffusible 13. VEGF165 is a 38 kDa disulphide-linked homodimer protein, and a potent and specific endothelial cell mitogen involved in the induction of angiogenesis, that is the growth of new blood vessels 14. VEGF165 binds the semaphorin receptor, Neuropilin-1 and promotes complex formation with VEGF R2 15. In this study, VEGF165 was selected because of the mentioned superiorities. VEGF protein has been isolated from HeLa cells 16, but, in this study, RNA was extracted and VEGF121 and VEGF165 gene from this cell was amplified for the first time.
Given the choice of parameters of primer designing, such as cost effectiveness and saving the time, the primers were designed in such a way that it could eliminate the sub-cloning step into T vector, by direct cloning of the gene in the expression vector 17. In the usual gene cloning, the gene is cloned in a T vector, digested by restriction enzyme and sub cloned in an expression vector for correct restriction site formation. The first stage (T vector cloning) was eliminated by addition of 2 nucleotides for BamHI and 3 nucleotides for XhoI on either side of their recognition site for an enzyme to cleave efficiently. So, the VEGF gene was cloned directly in the expression vector.
In this study, the isoform VEGF165 was chosen to be prokaryoticly expressed to soluble form. VEGF165_pET-32a(+) was expressed as Hisx6 and thioredoxin (trx) tagged protein. The 6Hisx helps to readily purify the protein with Ni-NTA affinity chromatography and the trx.tag as a highly soluble polypeptide can potentially enhance the expression and solubility of VEGF165 protein in E. coli 18,19. VEGF165 was cloned in pET-32a(+) vector as studied earlier 20 with some modifications and optimization of expression. The VEGF165 has been produced in various vectors (pET-22b, pET-28a, pET-30a, pQE30, etc…) and hosts by researchers 1,5,8,21,22,24 with a maximum expression level of 30% of the total protein. The expression of VEGF165 in pET-32a(+) was 40% of the total expressed protein which is significantly higher in comparison with the studies by other researchers.
The aim of this work was optimization of soluble VEGF165 expression in E. coli. In a study carried out by Bang et al the VEGF165 was expressed in a fix temperature of 37oC, while in this study, the temperature was optimized to get more soluble protein. The expression was optimized by adjustments such as induction time and incubation temperature for increasing the amount of soluble proteins 5. Parameters to increase the percentage of soluble protein were the temperature of 22oC and duration of 18 hr. Growth at 37oC resulted in accumulation of inclusion body while incubation at 22oC produced more soluble form of protein. In fact, this temperature helps the bacteria to consume all energy toward gradual protein biosynthesis and not cell division (Table 1) and thereby, lowering the rate of protein aggregation. This strategy facilitates the purification without denaturation, eliminating the problems such as urea purification process. Purification process at denaturing condition takes longer time with lower recovery and higher chance of miss folding 25-28. A variety of methods have been published describing refolding of insoluble proteins 28,29. Recombinant proteins expressed in E. coli are often produced as aggregates (Inclusion bodies). Even when inclusion bodies are formed, some proportion of the target protein is usually soluble within the cell. The high expression level of recombinant protein by the pET-32a(+) system limits the intracellular cell space and so it precipitates as aggregate forms. This is the key point in the importance of solubilization 30.
Conclusion :
In this study, the VEGF gene was isolated from HeLa cells for the first time and the expression vector for VEGF165 gene was constructed. Optimization of the expression parameters, i.e., time and temperature resulted in a high level of soluble protein expression in a prokaryotic expression system.
Acknowledgement :
The authors are grateful to Iranian Council for Stem Cell Sciences and Technologies-Vice president of science and technology for financial support (Grant NO: F-50). We express our thanks to our colleagues Dr. Mahboobeh Nazari, Dr. Saeed Talebi and Mr. Reza Hadavi for scientific cooperation.
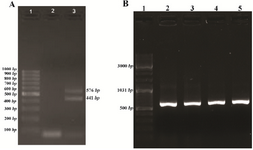
Figure 1. A) PCR amplification of VEGF165 gene from cDNA on agarose gel 1.5%. Lanes 1: 100 bp DNA ladder, 2: negative control and 3: PCR products of cDNA. B) PCR screening of the recombinant plasmid VEGF165_pET32a(+) on agarose gel 1.5%. Lanes 1: 100 bp DNA ladder, 2-5: PCR products.
|
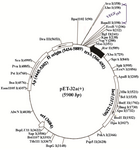
Figure 2. Schematic representation of recombinant expression vector. The VEGF165 gene inserted into the multiple cloning sites of pET-32a(+).
|

Figure 3. Sequence analysis of VEGF165. The BLAST result search for the highest homology found in GeneBank database.
|
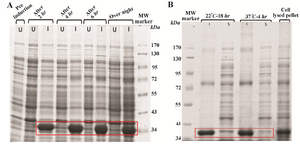
Figure 4. A) SDS–PAGE 12% (w/v) analysis of VEGF165 at different induction times. U=uninduced (-IPTG), I=induced (+IPTG). B) SDS-PAGE analysis of VEGF165. The effects of induction time and temperature on soluble and insoluble proteins expression. I=Insoluble. S=Soluble.
|
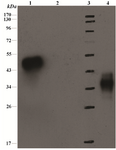
Figure 5. SDS-PAGE12% (w/v) analysis of VEGF165 and immunoblot with anti His Tag antibody. Lanes 1) positive control (His-Streptokinase), 2) negative control; bacterial lysate without insert, 3) protein molecular marker, 4) cell laysate of the transformed VEGF165_pET-32 a(+)construct.
|
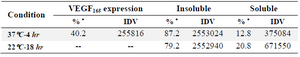
Table 1. Expression rate measured by AlphaEase FC Software. VEGF165 expressed 40.2% of the total protein (4 hr at 37°C). Insoluble: 87.2% of the total protein (4 hr at 37°C) and 79.2% (18 hr at 22°C). Soluble: 12.8% of the total protein (4 hr at 37°C) and 20.8% (18 hr at 22°C)
IDV=Integrated Density Value
* Based on integrated density value
|
|