Expression and Purification of Recombinant ROP1 of Toxoplasma gondii in Bacteria
-
Mohabati, Reyhaneh
-
Molecular Parasitology Laboratory, Department of Parasitology, Pasteur Institute of Iran, Tehran, Iran
-
Department of Biochemistry, Science and Research Branch, Islamic Azad University, Fars, Iran
-
Babaie, Jalal
-
Molecular Parasitology Laboratory, Department of Parasitology, Pasteur Institute of Iran, Tehran, Iran
-
Amiri, Samira
-
Molecular Parasitology Laboratory, Department of Parasitology, Pasteur Institute of Iran, Tehran, Iran
-
Talebzadeh, Mohammad
-
Molecular Parasitology Laboratory, Department of Parasitology, Pasteur Institute of Iran, Tehran, Iran
-
Darbouy, Mojtaba
-
Department of Biochemistry, Science and Research Branch, Islamic Azad University, Fars, Iran
-
 Golkar, Majid
Molecular Parasitology Laboratory, Department of Parasitology, Pasteur Institute of Iran, Tehran, Iran, Tel: +98 21 66968855; E-mail: majid.golkar@gmail.com
Golkar, Majid
Molecular Parasitology Laboratory, Department of Parasitology, Pasteur Institute of Iran, Tehran, Iran, Tel: +98 21 66968855; E-mail: majid.golkar@gmail.com
-
Molecular Parasitology Laboratory, Department of Parasitology, Pasteur Institute of Iran, Tehran, Iran
Abstract: Background: Toxoplasmosis is a worldwide-distributed infection which is mostly asymptomatic but can cause serious health problems in congenitally-infected newborns and immunecompromised individuals. Research is undergoing both to improve Toxoplasma serological tests, which play the main role in laboratory diagnosis of the infection, and develop an effective vaccine to prevent the infection. Some studies showed usefulness of rhoptry protein 1 (ROP1) antigen of Toxoplasma gondii (T. gondii) in serodiagnosis of the infection and induction of protective immunity. The purpose of this study was to produce recombinant ROP1 and evaluate its antigenicity against human infected sera.
Methods: DNA encoding ROP1, amino acids 171 to 574, was obtained from T. gondii RH strain by polymerase chain reaction amplification and cloned in prokaryotic expression plasmid pET-15b. rROP1 was expressed in Escherichia coli (E. coli) and purified in a single step by immobilized metal ion affinity chromatography.
Results: DNA sequencing showed 99% similarity between the cloned sequence and the corresponding sequence in Gene bank. Results indicated the proper antigenicity of rROP1. Sera from Toxoplasma infected individuals specifically recognized rROP1 in Western blotting.
Conclusion: rROP1 is antigenic toward human infected sera and can be used in studies for development of both a Toxoplasma serological test and a protective vaccine.
Introduction :
T. gondii is an obligate intracellular pathogen that can probably infect all warm-blooded animals including humans 1-3. Primary infection with T. gondii acquired during pregnancy can spread to the fetus and results in premature birth, permanent neurological damage and visual impairment. Although toxoplasmosis is generally benign in immunocompetent individuals, it is a significant cause of morbidity and mortality in congenitally-infected infants and immunocompromised individuals such as those infected with human immunodeficiency virus (HIV) or those receiving immunosuppressive therapy 3-5. Some studies reported if primary maternal Toxoplasma infection is diagnosed promptly, chemotherapy can reduce both transmission of the parasite to offspring and morbidity and mortality associated with the congenital infection 4-6.
Generally, laboratory diagnosis of Toxoplasma infection can be established by isolation of the parasite from blood or body fluids, demonstration of the parasite in tissues, detection of specific nucleic acid sequences with DNA probes, or detection of T. gondii-specific immunoglobulins 7. Diagnosis of toxoplasmosis in immunocompetent individuals e.g. pregnant women is mainly based on serological tests since more than 90% of pregnant women present no specific clinical signs of the infection 8. On the other hand, definitive diagnosis in immunocompromised individuals is mostly undertaken by direct detection of the parasite by means of PCR, histology, cell culture and mouse inoculation 9.
Serological tests are primarily based on crude Toxoplasma antigens which are quite laborious and expensive to produce. Moreover, the complex nature of crude Toxoplasma antigens makes standardization of the assays difficult and frequently gives rise to insufficient specificities for differentiation between acute and chronic infection 10,11. Recombinant antigens have been considered as an alternative for the development of improved serological tests 7,9,12,13.
The invasive stages of apicomplexans are characterized by the presence of an apical complex composed of specialized cytoskeletal and secretory organelles, including Rhoptries 14. Rhoptry proteins have key biological roles in penetration into the host cell, and may facilitate the formation of the Parasitophorous Vacuole (PV) and mediate its clustering with host cell organelles 15.
The ROP1 gene is a single copy gene with a message of approximately 2.1 kb (amino acids 1-667) and shows an unusual charge and amino acid asymmetry 16,17. Soluble ROP1 is secreted into the PV during entry of T. gondii into host cells before its quick disappearance. This rapid disappearance suggests that ROP1 plays a role in early invasion 18-20.
Diagnostic performance of ROP1 has been investigated in IgG, IgG avidity and IgM ELISA for detection of Toxoplasma infection and distinguishing between acute and chronic infection 5,21,22. Also, ROP1 has been assessed as a vaccine candidate against T. gondii infection in sheep and mice 23,24.
In this study, our purpose was to evaluate bacterial expression and purification of recombinant ROP1 protein (rROP1). The purified rROP1 presented proper antigenicity and could be used in future studies for exploring its usefulness in developing a recombinant ELISA test and/or effective vaccine against toxoplasmosis.
Materials and Methods :
Parasitic and bacterial strains: T. gondii RH strain was used for extraction of genomic DNA. Tachyzoites were injected into peritoneal cavity of Swiss mice. Three days later, tachyzoites were harvested from peritoneal fluid, washed with phosphate-buffered saline (PBS) and stored at - 80ºC until they were ready for use. E. coli Top 10F’ (Invitrogen, Carlsbad, CA, USA), E. coli BL21(DE3)PlysS and Rosetta (DE3) (Promega, Madison, WI, USA) were used for cloning and expression of recombinant antigen, respectively.
Construction of the recombinant expression plasmid pET15-ROP1: DNA sequence of rop1 was obtained from patent database (accession no. AAq77715.1). Genomic DNA was extracted from tachyzoites of T. gondii, RH strain, by a genomic DNA extraction kit (Bioneer, Seoul, South Korea) and used as the template for polymerase chain amplification (PCR) of ROP1 gene fragment, encoding amino acids 171 to 574 (404 aa) using the following primers:
Forward primer: ATTCATATGGCCCTTTC GAGCCACAATGG
Reverse primer: ATTCTCGAGCGATCCAT CATCCTGCTCTCTTCC
PCR amplification was performed as 1 cycle of 95ºC for 5 min then 40 cycles of 95ºC for 50 s, 59.8ºC (+0.1ºC increment per cycle) for 40 s and 72ºC for 50 s. The last step was a final primer extension for 30 min at 72ºC. The PCR product was ligated into T/A cloning vector, pTZ57R/T (Fermentas, Vilnious, Lithuania). Ligation product was transformed into E. coli Top 10F’ cells and recombinant clones were selected by blue/white screening, and a recombinant clone harboring ROP1 DNA was recognized by colony-PCR. The ROP1 DNA fragment was cut with XhoI and NdeI and inserted in-frame into expression vector pET-15b(+) which was previously cut with the same enzymes. The presence of ROP1 insert in the recombinant plasmid was checked by restriction analysis and sequence determination. The recombinant plasmid was named pET15b-ROP1.
Expression of recombinant ROP1 (rROP1): E. coli Rosetta (DE3) and BL21 (DE3) pLysS bacteria were transformed by pET15-ROP1and grown in Luria Bertani (LB) broth supplemented with ampicillin (100 µg/ml) and chloramphenicol (34 µg/ml). The culture was grown with vigorous shaking at 37ºC to an OD600 of 0.6-0.8. Protein production was induced by 0.1, 0.5, 1.0, and 1.5 mM isopropyl-β-D-thiogalactoside (IPTG), and bacteria were incubated by vigorous shaking for 4, 5, 6 and 16 hr in order to find the best expression conditions. Induced bacteria were harvested, lyzed and expression of rROP1 was analyzed by SDS-polyacrylamide gel-electrophoresis (SDS-PAGE). Expression of rROP1 was estimated by densitometry analysis.
Purification of rROP1: Bacteria from 500 ml induced culture were resuspended in 20 ml of buffer A (0.5 M NaCl, 30 mM Tris-base, pH=8.5 and 1 mM Phenylmethylsulfonyl Fluoride (PMSF) as a protease inhibitor). Cells were sonicated for 10 min using MSE ultrasonic disintegrator (Fisons, Loughborough, UK) at 60% power and centrifuged at 8500 rpm and 4ºC for 20 min. The supernatant was passed through a 0.2 µm (pore size) filter. Five ml of Ni2+-nickel Nitrilotriacetic Acid (NTA) resin, previously equilibrated with buffer A, was incubated with the supernatant and stirred gently for 1 hr at room temperature. Then, the resin was washed sequentially with 20 ml of buffers B, C and D (buffer A containing 20, 40, and 80 mM imidazole, respectively). rROP1 was eluted with buffer E (buffer A containing 400 mM imidazole) and dialyzed against PBS overnight at 4C.
Electrophoresis and western blot analysis: Protein samples were analyzed by SDS-PAGE using 12% polyacrylamide gel. Protein bands were transferred onto PVDF membrane. Membrane was blocked with 2% bovine serum albumin in PBS-0.05% Tween 20 and probed with 1:100 diluted pooled human sera representative of acute, chronic and non-infected pregnant women. Bound antibodies were detected using horseradish peroxidase conjugated goat anti-human IgG antibodies diluted 1:7000 (Sigma, Hilden, Germany). Human sera and secondary antibodies were diluted in blocking buffer. Signals were detected using 3, 3’- diaminobenzidine tetrahydrochloride substrate (DAB) (Kementec Diagnostic, Taastrup, Denmark).
Results :
Cloning of ROP1 gene fragment: A 1234 bp gene fragment encoding amino acids 171 to 574 (accession no. AAq77715.1) of ROP1 was PCR amplified from the genome of T. gondii RH tachyzoites and cloned into pTZ57R/T. The recombinant clone was selected by colony PCR and sub-cloned into pET-15b(+) expression vector. Sequence analysis of ROP1 gene fragment demonstrated 99% sequence similarity with the corresponding published sequence of rop1. In pET15-ROP1, 22 amino acids including a cluster of 6 histidine residues were fused to the N-terminal of rROP1 to facilitate subsequent purification of the recombinant protein.
Expression and purification of rROP1: The recombinant plasmid pET15-ROP1 was transformed into two different strains of E. coli i.e. Rosetta (DE3) and BL21 (DE3) pLysS. Expression of rROP1 was induced in both strains at different concentrations of IPTG. Moreover, a time-course study was performed to find out the optimized time of induction. SDS-PAGE analysis showed the presence of a protein band of about 80 kDa in induced bacteria which was absent in uninduced bacteria (Figure 1A). According to the densitometry analysis, rROP1 comprised 1.5% of total proteins in Rosetta (DE3) bacteria, while the expression level was lower in BL21 (DE3) pLysS bacteria and contributed to 0.5% of the total proteins (Figure 1A). We applied Rosetta (DE3) for further analysis. The expression of rROP1 was approximately the same when IPTG was added to the culture medium at increasing concentration from 0.1 to 1.5 mM (Figure 1B). The time-course study showed that the maximum expression of rROP1 was achieved 4 hr after induction (Figure 1C).
The apparent molecular weight of rROP1, about 80 kDa, was much higher than the molecular mass. It was probably due to the octa peptide repeats in ROP1 sequence which are rich in proline-glutamic acid residues 25.
Induced bacteria from a 500 ml culture were harvested and lysed by sonication. Soluble proteins were applied to Ni-NTA and rROP1 was purified in a single step. About 1.5 mg of rROP1 was obtained with the purity of 91% measured by Image J software (Figure 1D).
Immunoreactivity of rROP1 against human sera: To evaluate antigenicity of rROP1, 1 µg of the purified rROP1 was transferred onto a PVDF membrane and probed with pooled human sera representative of acute and chronic Toxoplasma infection. The result of immunoblotting showed both acute sera and chronic sera recognized rROP1 with approximately the same intensity (Figure 2).
Discussion :
In this study, we report expression and purification of T. gondii rhoptry protein 1 (ROP1) in E. coli as a soluble protein. Antigenicity of the purified protein was confirmed in immunoblotting using T. gondii-positive human sera.
In order to avoid interference of large fusion tags in ELISA and vaccination experiments, a short N-terminal fusion tag was used for production of rROP1. Though the use of E. coli Rosetta (DE3) host, which has been genetically engineered to increase expression of proteins with rare codons, increased the expression of rROP1, the protein was still poorly expressed. The concentration of IPTG could influence the expression of recombinant proteins. We found that IPTG concentration higher than 0.1 mM didn’t increase the expression of rROP1. Some studies showed the optimized expression of recombinant proteins in flask culture which can be achieved at IPTG concentration as low as 0.1 mM 26. The highest level of expression of rROP1 was achieved at 4 hr post induction and the protein yield was decreased afterward, possibly due to protein degradation by proteases. The presence of a 6 Histidine tag allowed purification of rROP1 by immobilized metal ion chromatography (IMAC). Single-step purification of rROP1 yielded highly pure protein. Western blot analysis showed specific immunoreactivity of rROP1 in sera from pregnant women with acute Toxoplasma infection, as well as women with chronic Toxoplasma infection.
Some studies reported higher production yield of ROP1, compared to the present study. However, they either expressed shorter length of the protein 27 or used large fusion partners 23. As there are 62 E. coli rare codons in ROP1 gene, chemical synthesis of a codon-optimized ROP1 gene might improve the expression level.
ROP1 has been used in IgM ELISA for the detection of acute Toxoplasma infection with very promising results 5,22. In fact, it was used in the development of a recombinant IgM ELISA test kit by Mikrogen Co. Recently, usefulness of ROP1 in avidity ELISA for discriminating acute from chronic infection was studied 21.
Several studies showed DNA vaccines encoding ROP1 protein induced strong protective immunity in animals 24,28. To our knowledge, no study used recombinant ROP1 protein in vaccination experiments, probably due to the difficulties in producing large amounts of the protein. Immunization of animals with recombinant ROP1 formulated in T-cell adjuvants or application of prime-boost strategy using a ROP1 DNA vaccine and recombinant ROP1 might improve the protective immunity.
Conclusion :
In conclusion, this study reported production of rROP1 of Toxoplasma gondii in E. coli. The recombinant protein was highly purified in a single chromatographic step and shown to be immnuoreactive toward sera from infected individuals. The purified protein can be used, in combination with other Toxoplasma antigens for developing ELISA tests, and in vaccination experiments for development of the protective immunity in animal models.
Acknowledgement :
This work was supported by the research grants No. 461 from Pasteur Institute of Iran and No. 8900846 from Iranian National Science Foundation.
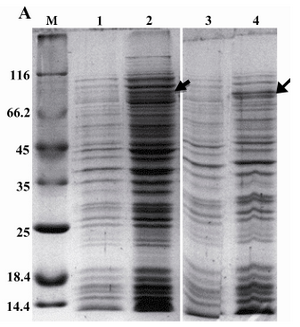
Figure 1A. Expression of rROP1 in two different bacterial expression hosts, E. coli BL21 (DE3) pLysS (lanes 1 and 2) and Rosetta (lanes 3 and 4). Induced (lanes 2 and 4) and uninduced bacteria (lanes 1 and 3) were analyzed on SDS-PAGE. Densitometry analysis using Image J software showed rROP1band attributed to 1.5% and 0.5% of total protein in induced Rosetta (DE3) and BL21 (DE3) pLysS bacteria, respectively
|
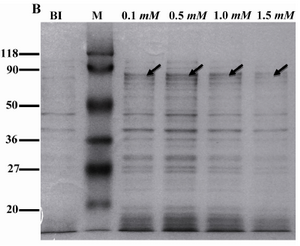
Figure 1B. Expression of rROP1 at different concentrations of IPTG. Rosetta (DE3) bacteria were induced with IPTG at concentrations from 0.1 mM to 1.5 mM. Densitometry analysis of rROP1 protein band showed no difference in protein expression when IPTG increased from 0.1 mM to 1.5 mM. BI: before induction
|
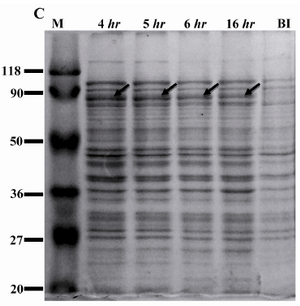
Figure 1C. Expression of rROP1 at different times of induction. Rosetta (DE3) bacteria were induced with 0.1 mM IPTG and incubated for different periods of time. Densitometry analysis of rROP1 protein band showed highest expression of rROP1 and was achieved 4 hr after induction.
BI: before induction
|
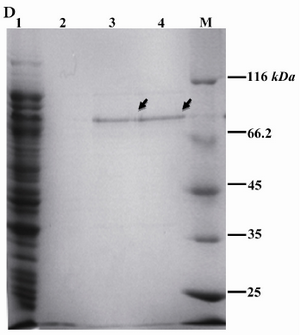
Figure 1D. Purification of rROP1. Soluble recombinant rROP1 protein was purified by Ni+2-NTA affinity chromatography, as illustrated in material and methods section, and analyzed by SDS-PAGE. Lane 1: flow through; lane 2: Wash; lane 3: Elution 1; lane 4: Elution 2
|
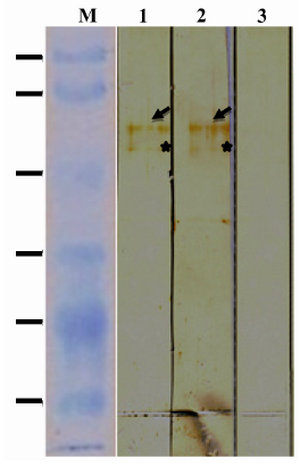
Figure 2. Immunoreactivity of rROP1. Purified rROP1 was analyzed by SDS-PAGE, transferred onto PVDF membrane and probed with pooled sera from pregnant women with acute (lane 1), chronic (lane 2) and no T. gondii infection (lane 3).
*Degraded ROP1 is marked with star
|
|