Overexpression and Purification of C-terminal Fragment of the Passenger Domain of Hap Protein from Nontypeable Haemophilus influenzae in a Highly Optimized Escherichia coli Expression System
-
 Tabatabaee, Akram
Department of Biology, Science and Research Branch, Islamic Azad University, Tehran, Iran, Tel: +98 21 64432432; Email: Akram_tabatabaee@yahoo.com
Tabatabaee, Akram
Department of Biology, Science and Research Branch, Islamic Azad University, Tehran, Iran, Tel: +98 21 64432432; Email: Akram_tabatabaee@yahoo.com
-
Department of Biology, Science and Research Branch, Islamic Azad University, Tehran, Iran
-
Yavari, Neda
-
Department of Microbiology, Karaj Islamic Azad University, Alborz, Iran
Abstract: Background: Nontypeable Haemophilus influenzae (NTHi) is a common cause of respiratory tract disease and initiates infection by colonization in nasopharynx. The Haemophilus influenzae (H. influenzae) Hap adhesin is an auto transporter protein that promotes initial interaction with human epithelial cells. Hap protein contains a 110 kDa internal passenger domain called “HapS” and a 45 kDa C-terminal translocator domain called “Hapβ”. Hap adhesive activity has been recently reported to be connected to its Cell Binding Domain (CBD) which resides within the 311 C-terminal residues of the internal passenger domain of the protein. Furthermore, immunization with this CBD protein has been shown to prevent bacterial nasopharynx colonization in animal models.
Methods: To provide enough amounts of pure HapS protein for vaccine studies, we sought to develop a highly optimized system to overexpress and purify the protein in large quantities. To this end, pET24a-cbd plasmid harboring cbd sequence from NTHi ATCC49766 was constructed and its expression was optimized by testing various expression parameters such as growth media, induction temperature, IPTG inducer concentration, induction stage and duration. SDS-PAGE and Western-blotting were used for protein analysis and confirmation and eventually the expressed protein was easily purified via immobilized metal affinity chromatography (IMAC) using Ni-NTA columns.
Results: The highest expression level of target protein was achieved when CBD expressing E. coli BL21 (DE3) cells were grown at 37C in 2xTY medium with 1.0 mM IPTG at mid-log phase (OD600 nm equal to 0.6) for 5 hrs. Amino acid sequence alignment of expressed CBD protein with 3 previously published CBD amino acid sequences were more than %97 identical and antigenicity plot analysis further revealed 9 antigenic domains which appeared to be well conserved among different analyzed CBD sequences.
Conclusion: Due to the presence of high similarity among CBD from NTHi ATCC49766 and other NTHi strains, CBD protein expressed here sounds to be theoretically ideal as a universal candidate for being used in vaccine studies against NTHi strains of various geographical areas. Further investigations to corroborate the potency of this protein as a vaccine candidate are under process.
Introduction :
Non-encapsulated Haemophilus influenzae or nontypeable Haemophilus influenzae is a
gram-negative bacterium that forms part of the normal flora of human nasopharynx. In addition, nontypeable Haemophilus influenzae (NTHi) is an important human respiratory tract pathogen that causes about 30% of otitis media in infants and children 1-3. Additionally, NTHi strains are intensified in adults suffering from chronic obstructive pulmonary diseases and likely implicated in the pathogenesis of the disease by chronically colonizing the lower respiratory tract 4-6.
Studies have shown that a critical step in the pathogenesis of NTHi disease is nasopharyngeal colonization 7. Bacterial surface adhesive structures, called adhesions, play a central role in NTHi colonization and finally pave the way for NTHi spread within the respiratory tract and even middle ear, sinuses, conjunctiva, or the lungs 8. One group of NTHi adhesins are auto transporters containing an N-terminal signal sequence, a C-terminal β-barrel domain and an internal passenger domain with effector function 9,10. The H. inflenzae Hap protein is a member of the large family of auto transporters and is present in all H. influenzae strains. Hap protein plays a crucial role in adhesion to respiratory cells, invasion, and bacterial aggregation 4, 11-13. Fink et al reported that, the internal passenger domain of Hap (HapS) harbors the adhesive responsibility, resides in 311 C-terminal amino acids of HapS and promotes the initial attachment of bacteria to respiratory epithelial cells.
Additionally, it has been found that the Cell Binding Domain (CBD) of HapS provokes immune response against NTHi colonization 14. One study revealed that immunization with CBD derived from three different strains led to a significant reduction in nasopharyngeal colonization. Therefore, the C-terminal fragment of HapS can be considered as a potential vaccine candidate 15. For exploiting HapS protein in vaccine research studies, availability of pure protein in large amounts is distinctly a need and this makes it necessary to try the best for optimizing a cost-effective expression/purification system that provides the required amounts of pure protein.
The main objective of this study is cloning and expression of CBD protein from H. influenzae ATCC49766 by a simple one-step procedure and presentation of optimized condition for obtaining large quantities of pure CBD for its application in vaccine studies.
Materials and Methods :
Bacterial strains and plasmids:Nontypeable Haemophilus influenzae ATCC49766 was obtained from the American Type Culture Collection and was grown at 37°C in brain heart infusion broth (Merck Company, Germany) supplemented with Hemin-L-Histidine (Sigma-Aldrich, Germany) and NAD (Mast Group Ltd, Liverpool, UK) 16. Escherichia coli (E. coli) strains DH5α and BL21 (DE3) and pET24a plasmid were provided by the national gene bank, Pasteur Institute of Iran.
Amplification of cbd gene:Genomic DNA of nontypeable H. influenzae ATCC49766 was extracted using Phenol-Chloroform extraction method as described by Sambrook et al 17. CBD of HapS (1000 bp) was PCR-amplified using HindIII-tailed forward (TAA GAA GCT TCG CGT TCA GAT TGG ACA GG) and XhoI-tailed reverse (ATA CTC GAG CAG GCT TTG ATC AGG CAG G), primer pairs and through the application of the following thermal program. It was conducted in 94°C for 7 min and followed by 35 cycles of 94°C for 1 min, 64.5ºC for 45 s, 72ºC for 1 min and a final extension time of 72ºC for 10 min.
PCR reaction (20 µl) contained 1X Pfu buffer with MgCl2, 1 mM dNTPs (Fermentas, St. Leon-Rot, Germany), 10 pmole of each primers, 1.25 U Pfu polymerase (Thermo, USA) and 100 ng of genomic DNA. The final product was purified by GeneJET™ Gel Extraction Kit (Fermentas, St. Leon-Rot, Germany).
Cloning and expression of cell binding domain of Haps protein:cbd PCR product was double digested by HindIII and XhoI and then ligated into pET24a, so that the C-terminus of recombinant protein could be expressed in fusion with 6xHis-tag. The integrity of the obtained construct, pET24a-cbd, was confirmed by restriction endonuclease analysis and bidirectional automated sequencing using BigDye reagent (Applied Biosystems, CA, USA) in Sequencing Center, Pasteur Institute of Iran. The nucleotide sequence of cbd gene fragment was deposited at the GenBank database under the accession number of KC261489.
To express the recombinant C-terminal fragment of HapS, pET24a-cbd recombinant plasmid was transformed into competent E. coli BL21 (DE3) cells and bacteria were grown on kanamycin-enriched (50 µg/ml) LB broth up to the optical density of 0.5. Following a 3 hr expression induction with 1.0 mM IPTG, the cell pellets were analyzed for protein expression, using 12% SDS-PAGE and CBD expression levels were determined using gel densitometry by Quantity One software (Bio-Rad), version 4.4.1.
CBD expression optimization:In order to express the highest possible level of CBD protein, several experimental parameters affecting the rate of recombinant protein expression including individual expressing single colonies, culture media (LB and 2xTY), induction stage (OD600 nm: 0.2, 0.4, 0.6, 0.8, and 1.0), inducer (IPTG) concentration (0.2, 0.5, and 1.0 mM), induction duration (2, 3, 5 hrs) and induction temperature (30, 35, and 37C) were analyzed by one factor at a time method. The protein expression levels in each experiment were quantified by scanning of SDS-PAGE gel and densitometric analysis of CBD protein band, as mentioned above.
Purification of 6xHis-tagged CBD protein:The best expression parameters obtained at the flask level (50 ml of bacterial culture) were scaled up to 1 L of culture. The pellet of E. coli BL21 cells expressing CBD potein was resuspended in lysis buffer (100 mM NaH2P04, 10 mM Tris-HCl, 8 M Urea, pH=8) by gentle vortexing and then centrifuged for 10 min at 11000 rpm. The obtained supernatant was loaded onto a nickel-nitrilotriacetic acid (Ni-NTA) column (Qiagen, Germany) for immobilized metal affinity chromatography (IMAC) purification under denaturing conditions based on the protocol provided by the manufacturer. After washing steps with wash buffer (100 mM NaH2P04, 10 mM Tris-HCl, 8 M Urea, pH=6.4), fractions of pure CBD protein were elued in optimized elution buffer (100 mM NaH2P04, 10 mM Tris-HCl, 8 M Urea, pH=4.5). Protein concentration was determined by Nanodrop analyzer (Thermo scientific, nanodrop1000 spectrophotometer) and the purity was determined by SDS-PAGE and Coomassie blue staining.
Western blot analysis:Proteins separated by SDS-PAGE were blotted onto a 0.45 µm pore size polyvinylidene difluoride (PVDF) membrane (Amersham Biosciences, Pittsburgh, USA) by using a semi-dry blotter unit (Bio-Rad Laboratories, California, USA). The membrane was probed, first with a Anti-6xHis rabbit antibody (Qiagen, Hilden, Germany), diluted to 1:1000 in TBS-Tween 20 and then with HRP-conjugated sheep anti-rabbit IgG (Sigma, Saint Louis, MO, USA) diluted to 1:3000 in PBS-Tween 20 for one hr at room temperature. Detection of CBD protein was achieved upon the development bands with DAB substrate (Sigma, Saint Louis, MO, USA) 18.
Sequence analysis of CBD protein:The amino acid sequences of three NTHi strains (N187, TN106, P860295) retrieved from GenBank were aligned with the sequence of CBD protein expressed in this study (ATCC49766) using Mega 4 software to assess the degree of amino acid conservation among them. An antigenicity plot of the studied CBD protein was also generated using the algorithm proposed by Kolaskar and Tongaonkar (http://imed.med. ucm.es/Tools/antigenic.pl) 19.
Results :
Construction of recombinant plasmids:The C-terminal fragment of HapS harbors the adhesive responsibility was amplified by PCR using specific primers. A 1% agarose gel was run to confirm the existence of proper size of PCR product and only a single band was appeared with an estimated size of ~1000 base pairs (Figure 1A). Then, the double digested PCR products and vectors were ligated together at a molar ratio of 3:1 and subsequently transformed into competent E. coli DH5α cells. The resultant colonies were evaluated for the true insert size by Colony PCR method. The oligonucleotide insertion into targeted plasmid was verified by double digestion of pET24a-cbd construct with two restriction enzymes, HindIII and XhoI simultaneously. Double digestion of correct oligonucleotide insertion should have produced two bands of 5300 bp, 1000 bp (Figures 1B and C).
Analysis and optimization of the expression of pET24a-cbd plasmid:Expression of genes linked to T7 promoters in E. coli BL21 (DE3) cells are triggered by adding IPTG. Expressed proteins in these cells along with uninduced ones were then separated using SDS-PAGE. After staining of proteins by Coomassie brilliant blue (R-250) and destaining, along with other bands, a band with an approximate size of ~37 kDa on lane of the induced samples was expected to be the protein of interest (Figure 2).
Six parameters of cultivation medium composition, growth phase at induction, IPTG concentration, the time of induction and incubation temperature after induction were modified in order to optimize the conditions for the production of recombinant CBD of HapS in E. coli BL21 (DE3) cells. To estimate the amounts of CBD production in each mentioned experiment, the resulting bands on SDS-PAGE gel were scanned and intensities of specific bands, corresponding to the protein of interest were measured.
Since freshly transformed colonies may differ significantly in their expression rates, CBD positive colonies were screened for high-level production of recombinant protein. Among seven CBD positive colonies, one colony showed a more intense band of target protein on SDS-PAGE gel and was selected for further investigations (Figure 3A). In media experiment, it was found that the maximum production of CBD occurred in 2xTY medium. On the other hand, protein band of interest on SDS-PAGE gel obtained from cultivation of E. coli BL21 (DE3) cells harboring pET24a-cbd in 2xTY/kanamycin was more intense in comparison with that obtained from LB medium/kanamycin. Furthermore, the ratio of the target protein band (CBD) to the density of total bacterial protein was higher in the case of 2xTY medium in comparison with LB (57% against 43%, respectively) (Figure 3B). This observation may be explained by the existence of higher amounts of yeast extract in 2xTY medium. As a result, the subsequent optimization experiments were conducted in 2xTY medium supplemented with appropriate antibiotics.
In addition to optimum starting culture, another important step for high-level protein production is determination about the appropriate growth phase at induction. For this purpose, different inoculation times of induction, OD600 nm: 0.2, 0.4, 0.6, 0.8, and 1 were considered. According to densitometric analysis, the suitable inoculation time of induction was at the mid-log phase (OD600 nm equal to 0.6) (Figure 3C). Thereafter, in order to examine the effect of inducer concentration on expression level of cbd gene fragment, different concentrations of IPTG, (0.2, 0.5, and 1.0 mM), were added to cultures at OD600 nm equals to 0.6. Intensity quantification of obtained target protein bands revealed that an increase in the IPTG concentration from 0.2 mM to 1.0 mM resulted in the improvement of cbd expression level. However, an increase in the IPTG concentration from 0.5 to 1.0 mM did not result in any significant improvement in recombinant protein production and the optimum IPTG concentration was considered 1.0 mM (Figure 3D).
In order to determine which time course of induction resulted in higher level of protein production, the same amount of IPTG, 1.0 mM, was added to the cultures at OD600 nm equals to 0.6 and at different times of 2, 3, 5 hrs post-induction, production levels of target protein was quantified. The optimal amount of CBD was attained over the period of 5 hrs after induction with IPTG (Figure 3E). The effect of different incubation temperatures (30, 35, and 37°C) on amount of protein production was also investigated under the optimum parameters obtained from the above experiments. Accordingly, 37°C was more suitable for protein expression following addition of IPTG (Figure 3F).
Purification and confirmation of expressed CBD protein:According to above experiments, the high expression level of CBD was obtained when CBD expressing E. coli BL21 (DE3) cells were grown at 37C in 2xTY supplemented with 50 μg/ml kanamycin and induction of cells were conducted with 1.0 mM IPTG after the OD 600 nm reached 0.6. The optimal expression level was achieved 5 hrs after the addition of IPTG to the medium. Therefore, the 6xHis-tagged C-terminal fragment of HapS was overproduced under the mentioned optimal conditions. Both supernatant and the pellet of cell lysates were checked for the presence of recombinant proteins and the majority of the expressed protein was detected in supernatant. Recombinant protein was purified with Ni-NTA affinity chromatography under denaturing conditions. Theoretical molecular weight of target protein was measured to be around ~ 37 kDa. The 37 kDa protein band was observed in SDS-PAGE and confirmed as CBD protein by western blot analysis using rabbit anti-His tagged antibody (Figures 4 and 5). The rate of CBD expression was approximately 62% among total bacterial proteins.
Sequence analysis of cell binding domain of HapS from different NTHi strains:Protein sequence alignment of the CBD of HapS from three NTHi strains N187, TN106, P860295 and CBD sequence employed in current study showed more than %97 identity (Figure 6A). The antigenicity plot of the CBD sequence from NTHi ATCC49766 indicated 9 antigenic domains that were similar to antigenic domains in three above strains. This finding propounds the possibility of using CBD (ATCC49766) as a universal antigen for vaccine studies in different geographical areas. The four prominent antigenic determinants were identified in the sequence also corresponded to the residues 13-20, 24-34, 199-210 and 230-239 (Figure 6B, Table 1).
Discussion :
Among auto transporter proteins of H. influenzae, Hap adhesin has been found to play a main role in initial bacterial colonization and aggregation. Since many surface exposed proteins of NTHi have found not to be conserved and they are considerably variable, based on Cutter et al’s studies, passenger domain of Hap (HapS) seems to be similar among different strains and immunization with it protects against colonization of NTHi in an animal model and subsequently prevents disease 5. Also, recent studies revealed that a part of passenger domain of Hap, the C-terminal fragment of HapS mediates bacterial interaction with host epithelial cells and contains CBD of Haps 20,21. Liu et al studied CBD from Haps from three strains of H. influenzae N187, TN106 and P860295. They reported that protein sequence alignment of CBD from mentioned strains showed 97.1% similarity and 81% identity. Also, they demonstrated that intranasal immunization with recombinant CBD elicits antibodies that cross-react with native HapS and finally concluded that C-terminal fragment of HapS protein (containing 311 residues) may be considered as a potential vaccine candidate against nontypeable H. influenzae 15. These results prompted us to investigate the immunogenicity of CBD from another strain of nontypeable H. influenzae and further determine whether it has any sequence homology with previously determined CBD protein sequences. Therefore, we constructed a recombinant C-terminal fragment of HapS protein from nontypeable H. influenzae ATCC49766, which is a routine strain for quality control of antibiotics and has been isolated from lung abscess 22.
In this work, pET24a-cbd harboring the cbd gene was constructed and its accuracy was confirmed by molecular biology and sequencing methods. Amino acid sequence alignment of CBD obtained in present study and previously published CBD amino acid sequences showed more than 97% identity and also its antigenicity plot identified 9 common highly antigenic domains that were located exactly in conserved regions among 4 different NTHi strains. These findings demonstrated that 311 C-terminal residues of HapS are highly conserved among NTHi strains as previously mentioned 8. Due to the presence of highly conserved antigenic epitopes among CBD from NTHi ATCC49766 and other NTHi strains, it is expected that the immune response against this protein could provide protection against different NTHi strains and develop the possibility for application of this protein as a universal target in vaccine studies.
For vaccine purposes, large quantities of pure protein are required, therefore, another specific aim of this study was directed towards establishing the condition to maximize the expression of CBD protein. Some factors which influence the yield of recombinant protein expression include host strain, expression vector, cell growth conditions, media composition, inducer concentration, time and duration of induction. The pET expression system in combination with the E. coli BL21 (DE3) cells is widely used for laboratory scale protein expressions. Cell growth conditions and media are important factors that affect protein expression levels; however, they seem to be protein-dependent and must be empirically determined in each specific case.
The induction conditions and post-induction time course are other important parameters that influence the production of the recombinant proteins and careful optimization is required to influence the productivity 23. In this work, the pET24a-cbd construct was transformed into E. coli BL21 (DE3) cells and the effect of different modifications in expression level including colony optimization, media composition, time of induction, IPTG concentration, and temperature and time course of induction was assessed. Eventually, the maximum expression level of CBD was obtained when CBD expressing E. coli BL21 (DE3) cells were grown at 37C in 2xTY supplemented with 50 μg/ml kanamycin and induction of cells was conducted with 1.0 mM IPTG after the OD600 nm reached 0.6. The optimal expression level was achieved 5 hr after the addition of IPTG to the medium. The ratio of the density of the target protein band (CBD) to that of total bacterial protein bands in induced cells was quantified to be around 62%. This level of expression was much higher than the one obtained in previous study by Liu et al, which reported the expression rate at most up to 43% of total bacterial protein 15. Notably, they reported the expression of cbd gene (three NTHi strains N187, TN106, P860295) under the control of T7 promoter (cloned into pET17b plasmid) in E. coli BL21 (DE3)/pLysS cells, which were grown at 37C in SOB broth medium supplemented with 1% glucose induction of cells was conducted with 1.0 mM IPTG at OD 600 nm: 1 to 1.5 for two hr.
Conclusion :
In our study, the overexpression was at least 2-fold higher than the one obtained by Liu et al 15. The optimization of expression parameters allowed us to increase the productivity of CBD protein as much as possible. Therefore, this study not only presents the optimized conditions for the production of large quantities of CBD, but also opens the gates for production of CBD-based conjugate vaccines like (Polyribose Ribitol Phosphate) PRP-CBD against both nontypeable and typeable H. influenzae. It is important to note that further investigation to test the in vivo immunogenicity and potency of the expressed CBD protein in animal models is in progress.
Acknowledgement :
This work was supported by Bacteriology and Hepatitis and AIDS Departments of Pasteur Institute of Iran.
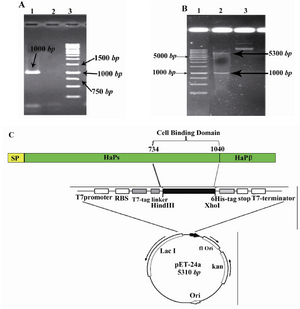
Figure 1. Cloning strategies for constructing the pET24a-cbd expression vector. A) Agarose gel electrophoresis of the PCR amplified fragment. From left to right, Lane 1: PCR product (shown with arrow), Lane 2: negative control, Lane3 : DNA Marker (Invitrogen, Karlsruhe, Germany); B) Restriction analysis of the pET24a-cbd construct. From left to right, Lane 1: DNA marker (Invitrogen, Karlsruhe, Germany), Lanes 2 and 3: digested and undigested forms of pET24a-cbd by HindIII/XhoI enzymes, respectively; C) Schematic representation of the expression elements in the pET24a-cbd plasmid; The cbd nucleotide sequence corresponding to amino acid residues of 734-1040 of NTHi strain P860295 was amplified by PCR and ligated into the HindIII/XhoI sites of the pET24a plasmid. This cloning strategy permitted to fuse T7 tag to the N-terminal and a 6xHis tag to the C-terminal of the CBD fragment. RBS and stop elements denote ribosme-binding site and translational stop codon, respectively
|
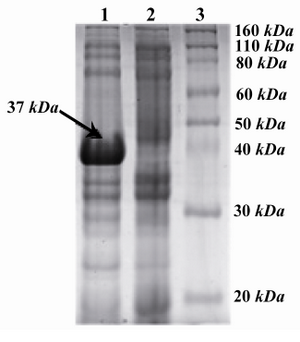
Figure 2. SDS-PAGE analysis of recombinant CBD with Coomassie-stained: From left to right, Lane 1: Induced with IPTG (arrow shows the ~37 kDa band), Lane 2: Non-induced with IPTG, lane 3: Protein marker
|
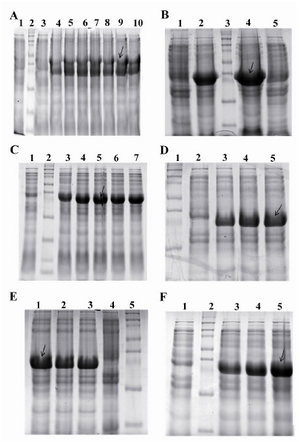
Figure 3. Optimization of the expression of pET24a-cbd plasmid. A) Colony optimization for high production level of CBD. From left to right, Lane 1: Uninduced with IPTG, Lane 2: Protein marker, lane 4-10: Induced with 1.0 mM IPTG (~38 kDa); B) Media optimization for high production level of CBD. From left to right, Lane 1: Uninduced with IPTG in LB broth, Lane 2: Induced with 1.0 mM IPTG in LB broth, lane 3: Protein marker, lane 4: induced with 1.0 mM IPTG in 2xTY, Lane 5: Uninduced with IPTG in 2xTY; C) Effect of inoculation time of induction on high production level of CBD. From left to right, Lane1: Uninduced with IPTG, Lane 2: Protein marker, Lane 3-5: induced with 1.0 mM IPTG at OD600 nm: 0.2, 0.4, 0.6, 0.8, and 1 respectively; D) Effect of IPTG concentration on high production level of CBD. From left to right, Lane 1: Protein marker, Lane 2: Uninduced with IPTG, Lane 3-5: induced with 0.2, 0.5, 1.0 mM IPTG, respectively; E) The effect of the period of induction on high production level of CBD. From left to right, lane 1-3: Induced with IPTG at time course of induction 2, 3, and 5 hr, respectively. Lane 4: Protein marker, Lane 5: Uninduced with IPTG; F) the effect of different incubation temperatures on high production level of CBD. From left to right, Lane 1: Uninduced with IPTG, Lane 2: Protein marker, Lane3-5: Induced with IPTG at 30, 35, and 37�C (the optimum result of each experiment was shown with arrows)
|
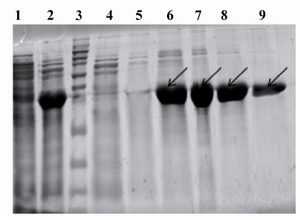
Figure 4. Purification of expressed CBD protein: From left to right, Lane 1: Uninduced with IPTG, Lane 2: Clear lysate, Lane 3: Protein marker, Lane 4: Flow through materials, Lane5: Wash fraction, pH=6.3, Lane 6-9: Elution fractions, pH=4.5 containing target protein (shown with arrows)
|
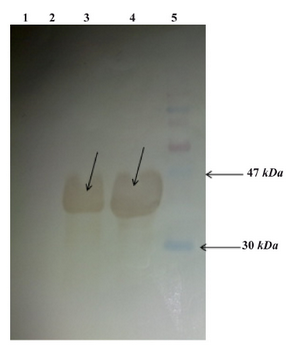
Figure 5. Western blot analysis: From left to right, Lane 1: E.coli BL21 (DE3) lysate, Lane 2: Uninduced E.coli BL21 (DE3) cells containing pET24a-cbd lysate, Lane 3: Induced E.coli BL21 (DE3) containing pET24a-cbd lysate, Lane4: Purified CBD, Lane5: Protein marker (shown with arrows)
|
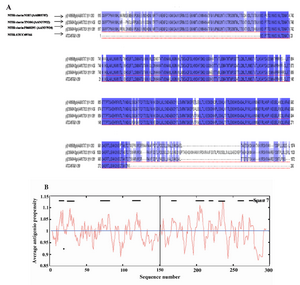
Figure 6. Sequence analysis of Cell Binding Domain of HapS from different NTHi strains. A) Sequence alignment of 3 different CBD amino acid reference sequences with the CBD sequence employed in this study (The fourth row). The CBD sequences were obtained from the NCBI gene bank database (accession numbers are shown). Identical and conserved amino acid residues are shown in black background and similar amino acids are shown in gray background; B) The CBD (amino acid 1�300) antigenicity plot of this study according to the Kolaskar and Tongaonkar method. The residues above the threshold level are hydrophilic. Threshold was set up for 8 residues
|
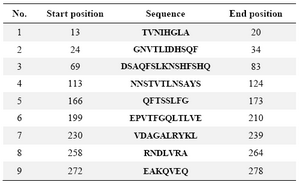
Table 1. The position of antigenic determinants of studied CBD
|
|