Culture of Ovine IVM/IVF Zygotes in Isolated Mouse Oviduct: Effect of Basal Medium
-
Farahavar, Abbas
-
Department of Animal Science, Faculty of Agriculture and Natural Resources, University of Tehran, Karaj, Iran
-
Shirazi, Abolfazl
-
Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Research Institute of Animal Embryo Technology, Shahrekord University , Shahrekord, Iran
-
 Khoram, Hamid
Department of Animal Science, Faculty of Agriculture and Natural Resources, University of Tehran, Karaj, Iran, +98 261 2248082; Hamid_kohram@yahoo.com
Khoram, Hamid
Department of Animal Science, Faculty of Agriculture and Natural Resources, University of Tehran, Karaj, Iran, +98 261 2248082; Hamid_kohram@yahoo.com
-
Department of Animal Science, Faculty of Agriculture and Natural Resources, University of Tehran, Karaj, Iran
-
Department of Clinical Sciences, Faculty of Veterinary Medicine, University of Shahid Chamran, Ahvaz, Iran
-
Zareh Shahneh, Ahmad
-
Department of Animal Science, Faculty of Agriculture and Natural Resources, University of Tehran, Karaj, Iran
-
Sarvari, Ali
-
Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Naderi, Mohammad Mehdi
-
Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Borjian Boroujeni, Sara
-
Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Zhandi, Mahdi
-
Department of Animal Science, Faculty of Agriculture and Natural Resources, University of Tehran, Karaj, Iran
Abstract: Background: The basal medium that supports Isolated Mouse Oviduct (IMO) is important for supporting embryo development and quality.
Methods: The culture of ovine IVM/IVF zygotes was done in IMO using SOFaaciBSA and SOFaaBSA as basal medium of IMO and in SOFaaBSA alone as control. For preparation of IMO mature inbred strain C57BL/6 female mice were synchronized and mated with vasectomized males. The females with vaginal plug were sacrificed and the zygotes were transferred in to the isolated oviduct at 20 hpi. The oviducts were cultured with SOFaaciBSA and SOFaaBSA for 6 days. Another group of zygotes were cultured in SOFaaBSA alone as control.
Results: Culture of zygotes in the IMO with SOFaaciBSA and SOFaaBSA, did not significantly affect the development and quality of embryos (p>0.05). The hatching rate, total and trophectoderm cells number in IMO groups’ blastocysts were significantly higher than SOFaaBSA alone. The morphological appearance of IMO blastocysts was superior to SOFaaBSA alone. When the quality of oocytes was poor, IMO could better support ovine embryo development either with SOFaaBSA or SOFaaciBSA than SOFaaBSA alone and there was a significant difference in blastocyst formation at day 6 with SOFaaBSA alone.
Conclusion: The culture of ovine IVM/IVF zygotes in IMO using two highly efficient ruminant embryo culture media not only could support development of ovine embryos similar to the level in non IMO culture system (SOFaaBSA alone) but also could improve the quality of resulting embryos. Additionally, IMO could better support the development of ovine embryos derived from poor quality oocytes compared to the SOFaaBSA alone.
Introduction :
The main factor affecting the quality of the in vitro produced blastocyst is the post fertilization culture conditions 1. IMO culture system has been used successfully in the culture of mouse, rat 2, hamster 3 and pig 4 and bovine 5,6 embryos from the one-cell to the morula/blastocyst stage. Basal medium that support IMO and cultured zygotes, is important for embryo development 6,7 and quality 5.
In conventional production of ovine embryos in vitro, the original formulation of SOF from Tervit et al (1972) with some modification (SOFaaBSA) was used for high rate of blastocyst production. SOFaaBSA is a glucose containing medium but viability of isolated mouse oviduct and cultured embryos with SOFaaBSA is unclear.
The culture of ovine zygotes in IMO with two highly efficient ruminant embryo culture media (SOFaaciBSA and SOFaaBSA), viability of explanted oviduct, cultured zygotes development and quality of resulted embryos have not been studied to date.
Material and Methods :
Except where otherwise indicated, all chemicals were obtained from the Sigma (St. Louis, MO, USA).
In vitro maturation and fertilization
Sheep ovaries were collected from slaughterhouse and the follicular fluid was collected in HEPES-buffered TCM199. The selected Cumulus Oocyte Complex (COCs) were matured for 24 hr in bicarbonate buffered TCM199 supplemented FBS, 0.1 iu/ml FSH and 0.4 mM sodium pyruvate. For IVF, matured oocytes were fertilized with Shall breed ram spermatozoa at concentration of 2×106 spermatozoa per ml. Semen from the same ram was used for all experiments.
Culture of zygotes in vitro: In vitro culture of zygotes was carried out in SOFaaBSA and used as control. The culture was done in a humidified atmosphere of 7% O2, 7.5% CO2, and 85.5% N2 at 39°C. Charcoal stripped fetal bovine serum (10% v/v) was added to each droplet 48 hr after start of culture (72 hpi).
Culture of zygotes in the isolated mouse oviduc: Zygotes were transferred to the mouse oviduct (approximately 5-7 per oviduct) at 20 hpi. The oviducts were transferred on to a Costar Transwell membrane (non-tissue culture treated, Nucleopore polycarbonate membrane, 3.0 µm pore size, 24 mm diameter, Costar 3414, Corning, NY). Approximately 1.5 ml of equilibrated medium had been placed in the compartment below the membrane insert. Plates were incubated at 39C under an atmosphere of 8% CO2 in air with maximum humidity. Embryos were recovered 6 days later (i.e., day 7 post insemination) by flushing with handling medium.
Mouse oviduct isolation: All animal experiments were performed in accordance with Avicenna Research Institute Ethical Committee Guidelines. For preparation of IMO mature inbred strain C57BL/6 female mice were synchronized and mated with vasectomized males. The females with vaginal plug were sacrificed and the zygotes were transferred in to the isolated oviduct at 20 hpi. The oviducts were cultured with SOFaaciBSA and SOFaaBSA for 6 days.
Embryo quality evaluation: Morphological evaluation: Embryo diameter at hatching was assessed by a microscopic grade. International Embryo Transfer Standards (IETS) were used for embryo evaluation by morphological methods. There are three grades of quality namely: excellent, good, fair and poor.
Embryonic cells: For differential staining of Inner Cell Mass (ICM) and Trophectoderm cells (TE) blastocysts were incubated in 0.2% Triton X-100 for 20 s and for staining of TE, immediately were transferred into 30 µg/ml Propidium Iodine (PI) solution for 1 min. For staining ICM were incubated in ethanol containing 10 µg/ml bisbenzimide (Hoechst) for 15 min on ice. Embryos were immediately mounted on glass slides and examined under an epifluorescence microscope.
Statistical analysis: Data analyses were carried out by one-way ANOVA using the SAS (Statistical Analysis System, 9.1 versions) software and GLM procedure. Shapiro-Wilk test was used for determining normality of data. A p-value of 0.05 by Tukey test and was considered significant. To present a true mean value of data the transformed means were reconverted. Moreover for representing the standard deviation, confidence limits of the transformed data were computed and then converted to linear scale.
Results :
Embryo development: This is the first study which showed that IMO could support ovine embryo development when cultured with two highly efficient ruminant embryo culture media (SOFaaciBSA and SOFaaBSA) as basal medium (Figure 1). There were no significant differences (p>0.05) in cleavage rate between experimental groups. Culture of zygotes in the IMO with SOFaaciBSA and SOFaaBSA, did not affect the proportion of zygotes development to the blastocyst stage at days 6, 7 and 8 compared to the respective control (p>0.05). There was a significant difference (p<0.05) between hatching rate of IMO group’s blastocysts compared with SOFaaBSA (Table 1). Results of this study showed that when the quality of oocytes are poor, IMO could support better ovine embryo development either with SOFaaBSA or SOFaaciBSA than SOFaaBSA alone (Table 2).
Embryo quality: Morphological evaluation: We generally had recovered more excellent embryos (51-57%) from the IMO (either with SOFaaciBSA or SOFaaBSA) and showing significant difference with SOFaaBSA alone (10.34%) (Figure 2, Table 3). The hatching of IMO embryos occurs at a smaller diameter (225±.42 and 240±8.1 vs. 257±6.42 m) than SOFaaBSA alone (Table 3). The IMO groups’ blastocysts had brighter appearance, with more prominent inner cell mass compared with blastocysts produced in SOFaaBSA alone (Figure 2).
Embryonic cells: There was no significant difference in the numbers of total embryonic cells and TE and ICM cells number of embryos cultured in IMO with SOFaaBSA. The numbers of total embryonic cells of embryos cultured in IMO with SOFaaciBSA and SOFaaBSA were significantly higher (p<0.05) than SOFaaBSA alone. The numbers of TE cells in IMO groups were significantly higher (p<0.05) than control. There was a tendency (p<0.05) to increase the number of ICM cells of IMO groups in comparison to control (Table 4). The ratio of ICM:TE did not show significant difference between groups. The normal allocation and integrity of inner cell mass between IMO produced blastocyst and SOFaaBSA alone after differential staining was significantly different (Figure 3).
Discussion :
Culture of ovine IVM/IVF zygotes was done in IMO using two highly efficient ruminant embryo culture media as basal media for support viability of the explanted mouse oviduct and cultured ovine embryos development and quality. SOFaaciBSA that was used as first basal medium of IMO was a glucose free medium which had been used by Holm et al (1999) for high bovine blastocyst development in vitro and supplemented with sodium citrate and myo-inositol. Citrate stimulates fatty acid synthesis 8 and is a chelator of metal ions (e.g., Ca2+), a feature that may be of importance for maintaining junctional integrity and thus of importance for compaction and blastocoel formation 9. However, it is possible that addition of citrate to SOFaaBSA further enhanced blastocyst development 10.
Myoinositol or its metabolites are essential components in cellular signalling and some metabolites may have a direct mitogenic effect 11. The embryotrophic properties of some batches of human serum have been correlated with high concentration of myoinositol 12. In the present study, there was no significant difference in blastocyst yield at days 6, 7 and 8 between zygotes cultured in IMO either with SOFaaciBSA or SOFaaBSA and SOFaaBSA alone (Table 1). It has been reported that the post fertilization culture environment, within certain limits, does not affect blastocyst development(1). The development of immature bovine and ovine oocytes to the blastocyst stage following maturation, fertilization and culture in vitro is generally limited to a frequency of about 40%. There is now a growing amount of evidence to suggest that while culture conditions during in vitro embryo production can impact the developmental potential of the early embryo, the intrinsic quality of the oocyte is the key factor determining the proportion of oocytes developing to the blastocyst stage 1,13. Results of our study were in agreement with published documents.
The hatching rate in our study was significantly different in IMO groups compared to the SOFaaBSA alone. In fact, 72-76% of the embryos cultured in IMO had been hatched from the zona pellucida by day 8, which was similar to the rate found in vivo 14. In our study culture of zygotes in IMO was done in 20% O2 atmosphere, while O2 percentage in conventional IVC (control) was 7% O2. It was reported that oxygen tension during IVC could negatively affect hatching rate 15. As a result, the higher hatching rate in IMO groups may be related to the other unknown beneficial effects of mouse oviduct on ovine embryo development in IMO.
Post fertilization embryo culture condition affects embryo morphology. It has been shown that presence of serum in embryo culture medium has provided materials linked to abnormal embryo morphology. The average diameter of blastocyst at hatching is significantly smaller for in vivo vs. in vitro derived blastocysts. The hatching of in vivo embryos at a smaller diameter could be indicative of an altered composition of zona pellucidae, causing a more rigid or elastic zona pellucidae and crack easily 16. Results of this study showed that hatching of IMO embryos occurs at a smaller diameter than SOFaaBSA alone and there were no significant difference between SOFaaciBSA and SOFaaBSA with IMO. In our study the morphological appearance of IMO produced ovine blastocysts (either in SOFaaBSA or SOFaaciBSA), was superior to those produced in SOFaaBSA alone. There was however no differences between IMO groups (Table 3). The IMO groups’ blastocysts had brighter appearance, more prominent inner cell mass and less irregularities compared with blastocysts produced in SOFaaBSA alone (Figure 1). It has been reported that embryos produced in defined or undefined SOFaaci medium are morphologically similar to the in vivo embryos, considering their lighter colour, tighter compaction, and the integrity of the inner cell mass 17. We did not observe morphologically differences between IMO produced embryos (Table 3). It is postulated that culture of ovine zygotes in IMO with SOFaaciBSA further improves the blastocysts quality due to the positive effects of myoinositol and citrate. These effects may unfold at ultrastructural level and gene expression patterns which are remained to be clarified.
The embryonic cell numbers, especially the ratio of ICM:TE cells, is one of the criteria for assessment of blastocyst quality 18. Aberrant allocation of ICM and TE cells in embryos at preimplantation stages may cause placental abnormalities and early fetal loss 19. The environment during early embryonic development clearly plays a crucial role in determining the cell numbers of the embryo. Small differences between in vitro and in vivo derived embryos already occur during development up to the 8-cell stage, but development in vivo during the first 100 hr post ovulation up to the fifth cell cycle is decisive for the total cell number of the embryo and the ICM in the expanded blastocyst at 7 days post ovulation 11,20. It was suggested that contact with the maternal tract might be of importance to switch on certain genes that encode for developmentally important processes, such as tight junction formation in the case of inner cell mass allocation 21.Therefore, it is speculated that in IMO groups, the embryonic cells are more stimulated to cleave because of their interaction with the oviductal cells. The embryonic cells number was not different between SOFaaciBSA and SOFaaBSA with IMO (Table 4). Therefore it can be concluded that SOFaaciBSA and SOFaaBSA can support ovine embryo cells division when cultured in IMO.
An aberrant ICM:TE ratio has been suggested to be related to the large offspring syndrome of in vitro-produced embryos 22. Therefore we did not observe significant differences in ICM:TE ratio between groups, but embryos cultured in IMO (either with SOFaaBSA or SOFaaciBSA) had approximately 2 fold higher total embryonic cells than SOFaaBSA alone.
Conclusion :
The culture of ovine IVM/IVF zygotes in IMO using two highly efficient ruminant embryo culture media not only supported development of ovine embryos as well as embryo culture in vitro alone but also the quality of resulted embryos improved. Animal preparation and synchronization cost and difficulty of embryo transfer may be some of the limitations of this work.
Acknowledgement :
We would like to thank Presidential Office Deputy of Science and Technology, Iran National Science Foundation (INSF) for financial support (Proposal No: 89003548).
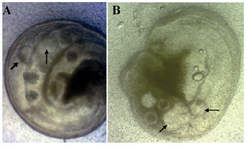
Figure 1. Ampulary segment of mouse oviduct containing ovine embryos 6 days after culture with, A) SOFaaciBSA; B) SOFaaBSA. Arrows show ovine blastocyst
|
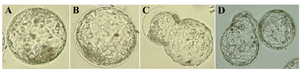
Figure 2. Morphological appearance of ovine blastocysts originated form, A) IMO with SOFaaciBSA; B & C) IMO with SOFaaBSA and D) SOFaaBSA alone. The integrity of the inner cell mass and more regular distribution of therphoctoderm cells in IMO produced embryos were considered during morphological evaluation
|
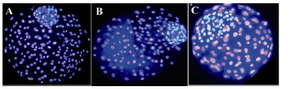
Figure 3. Differential staining of ovine blastocyst which had been originated from different culture conditions. A) IMO with SOFaaBSA; B) IMO with SOFaaciBSA; C) SOFaaBSA alone. The normal allocation and integrity of inner cell mass between IMO produced blastocyst and SOFaaBSA alone was significantly different
|

Table 1. Effect of post zygotic culture condition on ovine embryo development
*IMO: Isolated mouse oviduct.Values in the same columns with different superscripts differ significantly (p<0.05). Results were reported as Lsmeans±SD
** Hatching rate analyzed by chi-square test
|
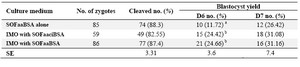
Table 2. Post fertilization development of poor quality ovine oocytes cultured in IMO
*IMO: Isolated mouse oviduct. Values in the same columns with different superscripts differ significantly (p<0.05). Results were reported as Lsmeans±SE
|
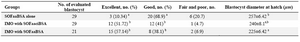
Table 3. Evaluation of blastocyst quality by morphologic methods in different post zygotic culture condition.
|
|