Radiotherapy Combination: Insight from Tumor Immune Microenvironment (TIME)
-
Alimohammadi , Masoumeh
-
Department of Immunology, School of Medicine, Tehran University of Medical Sciences, Tehran, Iran
-
Ghaffari-Nazari , Haniyeh
-
Department of Immunology, School of Medicine, Mashhad University of Medical Sciences, Mashhad, Iran
-
Alimohammadi, Reza
-
Department of Immunology, School of Medicine, Shahid Beheshti University of Medical Sciences, Tehran, Iran
-
Bakhshandeh, Mohsen
-
Department of Radiology Technology, Allied Medical Faculty, Shahid Beheshti University of Medical Sciences, Tehran, Iran
-
 Jalali , Seyed Amir
Department of Immunology, School of Medicine, Shahid Beheshti University of Medical Sciences, Tehran, Iran, E-mail: jalalia@sbmu.ac.ir, jalali5139@yahoo.com
Jalali , Seyed Amir
Department of Immunology, School of Medicine, Shahid Beheshti University of Medical Sciences, Tehran, Iran, E-mail: jalalia@sbmu.ac.ir, jalali5139@yahoo.com
-
Rezaei, Nima
Department of Immunology, School of Medicine, Tehran University of Medical Sciences, Tehran, Iran, Email: rezaei_nima@tums.ac.ir, rezaei_nima@yahoo.com
Abstract: The view of Radiotherapy (RT) as a simple inducer of DNA damage resulting in tumor cell death has dramatically changed in recent years, and it is now widely accepted that RT can trigger an immune response which provides a sound basis for combining RT with immunotherapy. Given that, radiation can be delivered with different regimens, its effect on immune responses and Tumor Immune Microenvironment (TIME) may vary with dose and fractionation schedule. This fractional dose dependency may need to be more considered because of recent developments in RT delivery techniques making it possible to deliver precisely higher dosages per fraction (hypofractionation) while reducing exposure to normal tissues. Although combining radiotherapy with immunotherapy could be a promising strategy for synergistic enhancement of treatment efficacy, the selection of the best-matched combination of immunotherapy with each radiotherapy scheme remains to be addressed. Thus, for designing better therapeutic combinations, it is necessary to understand the immunological effects of RT. Here, we review the impact of conventional and different hypofractionation radiation schedules on the TIME. Subsequently, we highlight how knowing about these interactions may have implications for choosing a rational combination with targeted therapies.
Introduction :
Since more than a century, radiotherapy has been a cornerstone of neoplastic burden management in patients with cancer. About 60% of patients with solid tumors receive Radiotherapy (RT) in the neoadjuvant, adjuvant, curative and palliative settings, 15% as monotherapy and 45% as radiochemotherapy, making it the most common treatment option for cancer 1,2. The first target of radiation within the tumor cells is the induction of double-strand breaks in DNA, leading to different forms of cell death and other cell cycle disturbances. Apoptosis, Mitotic Catastrophe (MC), autophagy, and senescence are consequences of radiation-induced DNA-damage 3.
It has been relatively common in radiation therapy research to focus on what radiation does to cancer cells and very little on what happens to the surrounding stroma, called Tumor Microenvironment (TME). A TME is both a structural and functional niche in which tumors progress. It is composed of cellular and molecular (cytokines, chemokines, extracellular matrix, and other molecules) parts. Its cellular components include tumor stromal cells [Cancer-Associated Fibroblast (CAFs), Mesenchymal Stromal Cells (MSCs), Endothelial Cells (ECs), pericytes] and a series of immune cells. Immune cells and their products form a microenvironment in tumor tissues called Tumor Immune Microenvironment (TIME) 4.
It is becoming increasingly clear that TME and thereby its immune subset, TIME, are changed dynamically in response to radiotherapy 5. The immunomodulatory effects of RT provide a compelling justification for combining RT and immunotherapy. Many studies indicate that the effects of RT on the TIME may vary with dose and fractionation schedule 6, thus in this review, we sought to delineate the TIME alteration in response to different RT regimens aiming to help to design more rational RT combinations by selecting immunotherapy targets which are fitting best to each RT scheme.
Types of radiotherapy techniques: There are two main types of radiotherapy to deliver radiation to the tumor location: internal radiation therapy (or brachytherapy) and external radiation therapy 7. Traditionally, the preferred method for obtaining the best therapeutic response in solid tumors is doses of 50 to 70 Gy delivered in multiple fractions of 1.8 to 2 Gy per fraction named conventional regimen 6. Recent technological developments in planning and delivery of external beam radiotherapy by using medical imaging before or even during treatment, have allowed more precise targeted delivery of larger doses of RT per fraction while minimizing dose to healthy tissues and organs. These approaches called hypofractionated and extreme hypofractionated radiation schedules, include Intensity-Modulated Radiation Therapy (IMRT), Image-Guided Radiation Therapy (IGRT), Stereotactic Radiosurgery (SRS), and Stereotactic Body Radiation Therapy (SBRT or stereotactic ablative radiotherapy - SABR) and have been used in the treatment of several tumors to augment tumor cell death while maintaining the therapeutic effectiveness. Using these techniques, single ablative doses as high as 20 to 24 Gy or extreme hypofractionated regimens such as 54 to 60 Gy in three fractions can be safely irradiated on small sites 8.
Radiotherapy in the immunotherapy era: Immunotherapy has been developed as a novel cancer treatment, over the past decade and has produced impressive outcomes; however, it has only benefited a small proportion of cancer patients 9. Thus, there is considerable demand for rational combination therapies that can augment its efficacy. Due to the strong immunostimulatory potential of radiotherapy, explained in the next part, it might be successful in making tumors more vulnerable to immune attack and thereby a rational option for combining with immunotherapy. Furthermore, there are some immunosuppressive effects of radiation that need to be overcome in order to take full advantage of anti-tumor immunity induced by radiotherapy and this can be obtained by combining with immunotherapy strategies that are targeting Immune-mediated suppressive mechanisms like immunosuppressive cells [such as Tregs, Myeloid-Derived Suppressor Cells (MDSCs), M2 macrophages] and inhibitory checkpoint molecules (so-called Immune Checkpoint Inhibitors or ICIs).
Since the advent of ICIs, the combination of RT with them has increasingly been used, and this combination overally has improved treatment outcomes in comparison to each therapy alone 10,11. However, more studies are needed to find out how the TIME changes induced by different radiation doses and fractionation may help ICI selection and affect treatment response.
Effect of RT on cancer cells: Besides direct or indirect DNA damage leading to cancer cell death, ionizing radiation also has a range of biological effects on cancer cells that those of which are deemed to trigger systemic immune response are discussed below. The effect of radiation on checkpoint expression on cancer cells is also discussed in the last part of this section.
Immunogenic cell death (ICD)-Through DNA damage induction and the following release of Damage-Associated Molecular Patterns (DAMPs), RT led to the ICD of tumor cells and transform them into an "in situ vaccine". The released antigens are phagocytosed by intra-tumoral cross-presenting Dendritic Cells (DCs) which will be matured and trafficked into the draining lymph nodes and subsequently activate naive T CD8+ cells. This RT-induced CTL response is mediated by DAMPs including ATP, High Mobility Group Box 1 (HMGB1), calreticulin and Heat Shock Proteins (HSPs), which are released following the ICD 12. ATP led to the recruitment of DCs to the tumor. Calreticulin on the surface of dying cells will be recognized by DCs and enhance phagocytosis. HMGB1 enables DCs to efficiently process and cross-present antigens by TLR4-dependent signaling and trigger a potent CTL response 13.
The cGAS-STING pathway: Ionizing radiation produces double-strand DNA breaks in cancer cells. DNA released following radiation-induced cell death has been found to activate the cyclic GMP-AMP synthase (cGAS)-Stimulator of Interferon Genes (STING) pathway, a pattern recognition receptor that senses cytosolic DNA 14. DNA binds to cGAS, which converts ATP and GTP to 2,3-cGAMP, a second messenger that activates the STING protein attached to the endoplasmic reticulum. STING activates interferon regulatory factor 3 (IRF3) that results in the production of type I interferons. A subtype of DCs (CD11c+CD8α+ BATF-lineage) specialized in antigen cross-presentation is thought to be the substrate of DNA-induced interferon production 15. Type I interferons induce cross-presentation of tumor antigens by DCs and subsequently lead to cross-priming of CD8+ effector T cells.
This induction of type 1 IFN by the cancer cells which is a critical mechanism for generating anti-tumor T cell response by RT, can be influenced by different radiotherapy regimens. Vanpouille-Box et al 16 demonstrated that DNA exonuclease Trex1 is a key regulator of radiation-induced immunogenicity. They found that this exonuclease is induced by radiation doses of more than 12–18 Gy in different mouse and human carcinoma cells, and degrades the RT-induced cytosolic DNA. Indeed, by increasing the radiation dose, more cytoplasmic dsDNA accumulates, until when Trex1 is apparently upregulated to degrade it. They reported the upregulation of Ifnb1 by RT regimen of 8GyX3, in contrast, 20 Gy radiation upregulated Trex1 in a patient-derived TP53/KRAS-mutated lung adenocarcinoma xenograft.
MHC class 1: The antitumor response of effector T cells includes recognition of tumor antigens and attack of cancer cells, which is related to MHC-I, the PD-1/PD-L1 axis, and the T cell receptor. There has been evidence for more than 20 years that radiation exposure may increase tumor cell MHC-I expression, which may enhance the immune recognition of tumors 17,18. Enhanced MHC-I expression in tumor tissues following SBRT increases the recognition of CD8+ T cells by in situ tumor-specific antigens. In early preclinical studies, the effects of hypofractionated radiotherapy regimens on the TIME are more robust than those of the conventional protocol, and the expression of MHC-I and related tumor peptides is higher with elevating radiotherapy dose 19. It has been reported that high doses improve tumor-specific CD8+ T cell infiltration by enhancing the upregulation of other immune signals 20. In addition to hypofractionated regimens, lower doses of radiation in conventional regimens have also been shown to upregulate MHC 21.
Checkpoint molecules: It has been shown that radiotherapy can affect the expression of checkpoint molecules in TIME. In a report on PD-L1 upregulation following RT, the authors believed that this effect is mostly due to inflammatory cytokines 22,23. Hiro Sato and colleagues conducted a study on how the signal of DNA damage regulates the expression of PD-L1 and showed that DNA Double-Strand Breaks (DSBs) lead to PD-L1 upregulation in a transcription-dependent way through DNA damage signaling molecules, including ataxia telangiectasia mutated and Ataxia Telangiectasia and Rad3-related (ATR), Chk1 and the downstream STAT-IRF1 pathway 24. In addition, they reported that the reduction of DNA repair proteins such as Ku80 and breast invasive carcinoma 2 (BRCA2) increased PD-L1 expression caused by DSBs. Their findings imply that RT may significantly increase PD-L1 expression in patients with these protein mutations; hence, PD-L1 blockade as a consolidation treatment after RT may be helpful in such cases. Furthermore, there is also some evidence in support of their findings 25,26. In a mouse model study, ATR inhibition decreased PD-L1 expression in cancer cells and improved T cell cell-killing ability 26 and in the combination of ATR inhibitor and radiation, a substantial delay in tumor development was found 25.
In another study, the mechanism of radiation-induced upregulation of immune checkpoint molecule expression has been reported related to the JAK2/STAT3 pathway 27. In this study both low-dose and regular-dose irradiation enhanced the expression of CD47 and PD-L1 molecules in lung cancer cells. It is noteworthy that, a recent data 28, demonstrated that the effect of RT on tumoral PD-L1 expression was regimen dependent, so that it was most durably with the fractionated regimen in comparison to their hypofractionated schemes.
The expression of PD-L1 on cancer cells' surface inhibits T cell activation by interacting with PD-1, which is the basis for using therapeutic antibodies to target it 29. However, a recent report has revealed another function of intracellular PD-L1 in stabilizing mRNA of DNA damage/repair proteins (like NBS1 and BRCA) 30. This discovery is fascinating as it suggests that intracellular PD-L1 may help cells survive radiotherapy, contributing to therapeutic resistance. Hence, blocking intracellular PD-L1 could be a useful strategy to improve the effectiveness of radiotherapy by impeding DNA repair.
Effects of RT on various immune cells: TIME in the TME includes a range of immune cell types that have various roles. Effector cells with cell-killing functions kill tumor cells through different mechanisms in both innate and adaptive immune responses. TIME immunosuppressive cells include MDSCs, CD4+ FOXP3+ regulatory T cells (Tregs), M2 macrophages, and some B-cell subsets 31. In the TME, lymphocytes are the most radiosensitive cells (Figure 1), followed by MDSCs, tumor-associated macrophages, and DC, which have higher radioresistance, respectively. Among the lymphoid population, Tregs are more radioresistant than the others, while NK cells and B lymphocytes are the most radiosensitive ones 32.
Lymphoid populations: It has been found that ablative or hypofractionated radiation activates the antitumor CD8+ T-cell response more effectively than conventional fractionation schemes 35-38. A single dose of 15 Gy radiation was compared to fractionated radiotherapy (5×3 Gy) by Lugade et al 39 and it was found that in comparison to fractionated schemes, the 20 Gy single schedule elevated Antigen-Presenting Cell (APCs) and CD8+ T cell numbers and enhanced the bioactivity of CD8+ T cells. In the Filatenkov study 40, an ablative single dose (30 Gy) induced a potent CTL response, while delivering a fractionated radiation (10×3 Gy) subsequent to the first dose diminished the response. Based on these results, it appears that conventional regimens may lead to the death of infiltrating lymphocytes, resulting in recurrences. In contrast, there are some supporting data on the optimal induction of CD8+ responses following fractionated radiotherapy 36,41. Dewan et al 36 compared three different radiotherapy schemes (1×20 Gy, 3×8 Gy, 5×6 Gy) in TSA breast carcinoma xenografts and demonstrated that the hypofractionated schedule of 3×8 Gy caused an abscopal effect in combination with CTLA-4 blockade. Furthermore, a conventional fractionation regimen (5×2 Gy) in combination with anti-PD-1 increased T-cell trafficking to locally treated tumor sites, being able to mediate the abscopal effect 41. Despite radiation's ability to stimulate antitumor immunity, some regimens have been demonstrated to enhance Treg numbers, hindering its therapeutic capacity.
In mice without tumors, whole-body radiation as low as 0.25 to 0.5 Gy, which is comparable to that used in conventional radiotherapy, has been found to increase Tregs 42. The same research also discovered that a local, single dose of radiation (10 Gy) administered to a mouse model of melanoma led to an increase in Tregs in the tumors and spleen. It has been demonstrated that local irradiation with 10, 20, or 30 Gy reduces CD8+ and CD4+ T cells but has no impact on FoxP3+ Tregs 43. The greater radioresistance of Tregs compared to other T-cell subtypes is assumed to be the cause of these increases in Tregs after radiation. Contrarily, FoxP3+CD4+ Tregs were reported to be diminished in quantity and to function less effectively after hypofractionated radiation therapy (5-8 Gy) in a mouse melanoma model 44. It is most probable that the radiation dosage and fractionation schedule will affect the variations in Treg enrichment or depletion. A local, single dosage of 5 Gy raised the Treg population but not the effector T-cell response in a mouse model of B16 murine melanoma expressing the antigen OVA 20. In contrast, a single radiation dosage of 7.5 or 10 Gy enhanced anti-OVA T cells, while decreasing the number of Tregs in the spleen, but a single radiation treatment of 15 Gy raised both the effector T-cell response and the Treg population. In this study, the Treg population was at its lowest with a 2x7.5 Gy schedule, which had the highest tumor control.
Myeloid populations: MDSCs are a diverse class of myeloid cells that are distinguished by their capacity to cause immunosuppression 45. They gather within the TME in response to the proinflammatory milieu and inhibit the activation of both CD4+ and CD8+ T-cell responses 46. Wu et al 47 demonstrated, employing a murine prostate cancer model, that a single high dose of radiation (15 Gy) induced the release of IL-6 from tumor cells and led to the recruitment of MDSCs. In a similar vein, tumors treated with radiation (5×3 Gy) in two prostate xenograft models—RM-1 and Myc-Cap—showed increases in M-MDSC and G-MDSC 48.
In addition to the above reports showing MDSC induction by radiation regardless of dose and fractionation, some reports are showing the fractional dose dependency of radiation effects on MDSCs 35. Treatment with Ablative Hypofractionated RT (AHFRT) 49 and a single high-dose scheme 40 led to the removal of MDSC, while treatment with conventional fractionated RT (CFRT) caused the accumulation of MDSC 50,51.
The polarization of macrophages is also affected differently depending on the dosage per fraction of RT. It has been demonstrated that conventional radiation doses shift TAMs toward the pro-inflammatory phenotype M1 MQ, which contributes to an anti-tumor immunity. Radiation therapy delivered as 4 Gy in 2 fractions caused tumor-infiltrating macrophages to change from M2 phenotype to M1 phenotype in pancreatic tumor xenografts 52. Similarly, in murine models of melanoma and pancreatic cancer, low-dose radiation treatment induced an increase in the infiltration of T cells into tumors as well as the killing of tumor cells through the production of TH1 cytokines by iNOS+ M1 macrophages 52,53. Additionally, several investigations have shown M2 polarization following a single high-dose and hypofractionated radiation scheme 54,55. M2 polarization frequently occurs in hypoxic regions, suggesting that high-dose radiation-induced vascular damage and hypoxia may encourage it 56.
Conclusion :
Understanding how SBRT, SRS and conventional fractionation affect the TME differently can help to design more effective combination therapies. Although single dose and hypofractionation regimens can increase CD8+ T cell responses, their deleterious effect on lymphocytes leading to Treg accumulation (given Tregs are more radioresistant than any other lymphocytes) and other TME changes like M2 polarization and enhanced tumor hypoxia may reduce their efficacy. Additionally, although conventional methods may not induce in situ immunization or T cell responses as potent as those produced by hypofractionation ones, they may reprogram macrophages toward the M1 phenotype, improving the anti-tumor response (Figure 2). Thus, it can be concluded that specific targeting of tumor-infiltrating Treg cells seems to be a more rational combination choice with hypofractionation RT while conventional RT, which recruits MDSCs into the TME, may need to be combined with MDSC-targeting therapy strategies. Furthermore, based on previously mentioned evidence, combining RT with ICIs (such as anti-PD-L1 and anti-CD47) are necessary for making the most of radiation-induced anti-tumor immunity.
In addition, it's important to note that a combination of both high-dose and fractionation regimens may augment the antitumor effects, which was first applied and named ‘RadScopal’ treatment by Barsoumian et al 57. Nevertheless, a debate still remains about the best combination method for each RT system and more studies are needed to validate the mentioned suggestions.
By expanding body of knowledge regarding the effects of radiation on the TME, new radiobiological models are required to enable the radiation oncologist to optimize the treatment plan based on the characteristics of the TME. Due to improvements in treatment delivery methods, hypofractionated treatments with dose per fraction greater than 10 Gy are becoming increasingly popular and accepted in radiation oncology; as a result, new radiobiological models are being created. Conventional fractionation models that use the LQ equation and the 4Rs of radiobiology—Repair of sublethal DNA damage, Redistribution in cell cycle, Repopulation, and Reoxygenation—originally only looked at the differences in radiation effects between normal and tumor cells but nowadays they have expanded to include other radiobiological processes as well 6. To account for the variations in response to RT owing to genetic heterogeneity amongst tumors of the same kind, the addition of a fifth R, Radiosensitivity, was proposed by G. Gordon Steele in 1989 58. Recently, two more Rs—Remodeling of the TME 59 and Rejection by the immune system 60—have been proposed to model the interactions between radiation and the tumor-associated stroma and the immune system. The practice of radiation oncology would reach a completely new level of combinatorial targeted and radiotherapies with the further incorporation of these additional Rs. Incorporation of these principles into treatment planning systems could enable precision treatment design for personalized medicine.
Acknowledgement :
This study was supported by a grant from Tehran University of Medical Sciences (grant n. 41391).
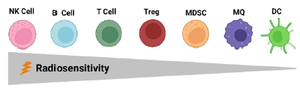
Figure 1. Radiosensitivity of tumor microenvironment immune cells. Within the lymphoid population, regulatory T cells (Tregs) are more radioresistant than any other T cells and B cells 33. NK cells and B lymphocytes are the most radiosensitive immune cells, while DCs are the most resistant ones 34.
|
![<p>Figure 2. Fractionation and dose can affect the immunomodulatory properties of radiotherapy. Depending on the fractionation regimen, RT can alter the expression of the immunotherapy targets through recruiting various immune cell types. Small doses of radiation delivered in conventional fractionated radiotherapy polarize macrophages to M1 phenotype, however, it also leads to MDSCs accumulation. Higher doses of radiation, such as those used in hypofractionation schemes, have a more potent immunostimulatory effect mediated by increased (APC) maturation as well as augmented T cell infiltration and enhanced expression of immunogenic proteins like MHC class I. This immunostimulation is also believed to be decreased by immunosuppressive mechanisms in the tumor microenvironment, like Tregs and M2 macrophages induced by these regimens. Once the radiation dose exceeds 12–18 Gy, immunogenicity is compromised because TREX1, an exonuclease, is activated, causing cytoplasmic DNA to degrade and negatively affect the cGAS–STING pathway which is necessary for radiation-induced immunogenicity. Furthermore, doses per fraction greater than 10 Gy may cause vascular damage thereby decreased vascular flow and hypoxia. [Treg: regulatory T Cell; MDSC: Myeloid-derived suppressor cells; PD-L1: Programmed-Death Ligand 1; DNA: Desoxyribonucleic Acid; MHC-1: Major Histocompatibility Complex class I; RT: Radiotherapy; T-REX: Three-Prime Repair Exonuclease].</p>](Images/Articles/60552/f2_small.png)
Figure 2. Fractionation and dose can affect the immunomodulatory properties of radiotherapy. Depending on the fractionation regimen, RT can alter the expression of the immunotherapy targets through recruiting various immune cell types. Small doses of radiation delivered in conventional fractionated radiotherapy polarize macrophages to M1 phenotype, however, it also leads to MDSCs accumulation. Higher doses of radiation, such as those used in hypofractionation schemes, have a more potent immunostimulatory effect mediated by increased (APC) maturation as well as augmented T cell infiltration and enhanced expression of immunogenic proteins like MHC class I. This immunostimulation is also believed to be decreased by immunosuppressive mechanisms in the tumor microenvironment, like Tregs and M2 macrophages induced by these regimens. Once the radiation dose exceeds 12–18 Gy, immunogenicity is compromised because TREX1, an exonuclease, is activated, causing cytoplasmic DNA to degrade and negatively affect the cGAS–STING pathway which is necessary for radiation-induced immunogenicity. Furthermore, doses per fraction greater than 10 Gy may cause vascular damage thereby decreased vascular flow and hypoxia. [Treg: regulatory T Cell; MDSC: Myeloid-derived suppressor cells; PD-L1: Programmed-Death Ligand 1; DNA: Desoxyribonucleic Acid; MHC-1: Major Histocompatibility Complex class I; RT: Radiotherapy; T-REX: Three-Prime Repair Exonuclease].
|
|