Application of Menstrual Blood Derived Stromal (stem) Cells Exert Greater Regenerative Potency Than Fibroblasts/Keratinocytes in Chronic Wounds of Diabetic Mice
-
Mirzadegan, Ebrahim
-
Nanobiotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Golshahi , Hannaneh
-
Nanobiotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Saffarian, Zahra
-
Nanobiotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Edalatkhah, Haleh
-
Nanobiotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Darzi, Maryam
-
Nanobiotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Khorasani, Somayeh
-
Nanobiotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran
-
Saliminejad, Kioomars
-
Reproductive Biotechnology Research Center, Avicenna Research Institute, ACECR,, Tehran, Iran
-
 Kazemnejad, Somaieh
Nanobiotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran, Tel: +98 21 22432020; Fax: +98 21 22432021; E-mail: s.kazemnejad@avicenna.ac.ir
Kazemnejad, Somaieh
Nanobiotechnology Research Center, Avicenna Research Institute, ACECR, Tehran, Iran, Tel: +98 21 22432020; Fax: +98 21 22432021; E-mail: s.kazemnejad@avicenna.ac.ir
Abstract: Background: In this study we differentially showed the effects of cell-seeded bilayer scaffold wound dressing in accelerating healing process in diabetic ulcers that still remains as a major clinical challenge. The aim of the study was to compare immunomodulatory and angiogenic activity, and regenerative effect differences between Menstrual blood-derived Stem Cells (MenSCs) and foreskin-derived keratinocytes/fibroblasts.
Methods: The streptozotocin-induced diabetic mice model was developed in male C57/BL6 mice. A bilayer scaffold was fabricated by electrospining silk fibroin nanofibers on human Amniotic Membrane (AM). Dermal fibroblasts and keratinocyte isolated from neonatal foreskin and MenSCs were isolated from the menstrual blood of healthy women. The diabetic mice were randomly divided into three groups including no treatment group, fibroblast/keratinocyte-seeded bilayer scaffold group (bSC+FK), and MenSCs-seeded bilayer scaffold group. The healing of full-thickness excisional wounds evaluations in the diabetic mice model in each group were evaluated at 3, 7, and 14 days after treatment.
Results: The gross and histological data sets significantly showed wound healing promotion via re-epithelialization and wound contraction along with enhanced regeneration in MenSCs-seeded bilayer scaffold group with the most similarity to adjacent intact tissue. Immunofluorescence staining of mouse skin depicted a descending trend of type III collagen along with the higher expression of involucrin as keratinocyte marker in the MenSCs-seeded bilayer nanofibrous scaffold group in comparison with other treatment groups from day 7 to day 14. Moreover, higher levels of CD31 and von Willebrand factor (VWF), and also a higher ratio of M2/M1 macrophages in association with higher levels of the neural marker were observed in the bSC+MenSCs group in comparison with bSC+FK and no treatment groups.
Conclusion: Healing symptoms in wounds dressed with keratinocyte/fibroblast-seeded bilayer scaffold was significantly lower than MenSCs-seeded bilayer scaffold done on impaired diabetic wound chronicity.
Introduction :
Worldwide increase in incidence of chronic non-healing diabetic wounds remain an ongoing challenge in advanced wound therapies. The prevalence of diabetes is increasing and expected to exceed 640 million people in 2040 1. Treatment of diabetic wounds face complexity and poor healing outcomes that lead to more than half of diabetic wound cases that do not heal expressively. Hence, multidisciplinary approaches are needed to treat these groups of patients 2.
Cell-loaded bioengineered skin graft substitutes open a new horizon in healing and regeneration of problematic wounds such as, large wounds, burns, pressure sores, diabetic ulcers, etc 3. Three major components of a bioartificial skin including, Extra Cellular Matrix (ECM), cells and growth factors are responsible for dynamic wound closure and wound resolution. In normal wound healing process, a cross talk between fibroblast and keratinocyte modulate inflammatory stage toward synthesis and remodeling phase. Fibroblasts are the most populated stromal cells in dermis 4,5 and begin to migrate into the wounded area and proliferate to synthesize various ECM components such as collagen, elastin, laminin, fibronectin, and glycosaminoglycans 6. Thus, the structural and mechanical integrity of sub-epithelial connective tissue is continuously maintained by fibroblasts. Furthermore, these cells have vital role in proliferation and migration of keratinocytes as well as modulation of the matrix metalloprotease activity based on a bilateral interplay with keratinocytes, which is critical during wound remodeling 7,8. During closing of the wound defect, keratinocytes proliferate and migrate over underlying granulation tissue towards each other to close the wound and restore the physical barrier. In a reciprocal manner, keratinocytes stimulate fibroblasts to produce and secret growth factors which in paracrine response regulate epidermal keratinocyte proliferation and turnover 9. Therefore, evaluation of fibroblast/keratinocyte-loaded bio-artificial skin grafts may provide new approach for regeneration of diabetic wound healing in comparison to mesenchymal-seeded bioengineered skin graft substitutes.
Furthermore, skin resident Mesenchymal Stem Cells (MSCs) are mainly located at the perivascular niche 10-14 and the granulation tissue at early stages after injury and become the most activated cells located in the granulation tissue. The MSCs proliferate and synthesize large amounts of the ECM, and upregulate growth factor expression which stimulates keratinocyte proliferation 15,16. In fact, activated mesenchymal cells via epithelial-mesenchymal 17 and keratinocyte-fibroblast interactions 9 regulate skin homeostasis during wound healing process.
In a previous experiment we fabricated a bilayer skin substitute consisting of human Amniotic Membrane (AM) which was coated by an electrospun silk fibroin with its related in vivo biological behavior was approved 18. In another study, we showed efficient wound healing using a synthetic nanofibrous bilayer skin substitute in a murine wound healing model 19. As shown in very recent research, we demonstrated the superior capability of Menstrual blood Stem Cells (MenSCs)-seeded bilayer scaffold wound dressing to improve and accelerate the regenerative process in diabetic wound bed in comparison to bilayer scaffold alone, and AM as novel biologic wound dressings 20. As an emerging concept, numerous investigations utilized MSCs from different sources due to their therapeutic properties including migratory, angiogenic, immunomodulatory and reparative potency 21-24. Among mesenchymal stem cells, MenSCs have been preferred due to their important practical advantages such as availability, easy accessibility and lacking ethical problems.
The purpose of the present study was to reproduce our very recent experiment comparing commercial cell-based tissue engineered skin substitutes that are usually derived from human keratinocytes and fibroblasts supported by a scaffold of synthetic mesh or xenogeneic collagen. Accordingly, in this study we used neonatal foreskin-derived fibroblasts and keratinocytes seeded on bilayer scaffold composed of human AM and electrospun silk fibroin nanofibers 18,19 to treat full-thickness wounds in diabetic mice model in comparison to MenSCs seeded on the same scaffold wound dressing.
Materials and Methods :
Ethics: This study was approved by bioethics committee of Avicenna Research Institute for all procedures and protocols (IR.ACECR.Avicenna.REC.1396.11). Forty-five ten-week-old male C57BL/6 mice weighing between 21-24 g were purchased from Pasteur Institute of Iran (Tehran, Iran). All animals utilized in this experiment, received humane care according to the Guide for the Care and Use of Laboratory Animals published by the National Institutes of Health 25. Human placentas were obtained from healthy mothers with no-complicated term pregnancies after elective caesarean birth at Iran-Mehr Hospital (Tehran, Iran). All women provided written informed consent. Menstrual Blood (MB) samples were collected from healthy volunteer women who signed written informed consent for use of their samples and experiment data. Keratinocytes and fibroblasts were isolated from neonatal foreskin tissue harvested from circumcised healthy newborns aged 2-10 months. Each donor’s parent signed an informed consent form and accepted by the Medical Ethics Committee of Avicenna Research Institute.
Isolation, cultivation, and characterization of MenSCs: MenSCs were isolated according to our protocol previously published 26. Briefly, MB was collected at the second day of menstruation by a sterile Diva cup (Diva International Co, Lunette, Finland). The collected MB was resuspended in Phosphate Buffered Saline (PBS) supplemented with 2.5 µg/ml fungizone (Gibco, Scotland, UK), 100 µg/ml streptomycin, and 100 U/ml penicillin (Sigma-Aldrich, MO, USA), as well as 0.5 mM ethylenediamine tetraacetic acid (EDTA) (Merck). Mononuclear cells were isolated from MB suspension by Ficoll-Hypaque density-gradient centrifugation (GE-Healthcare) and washed in the supplemented PBS. The cells were cultured in complete DMEM-12 (Gibco, Invitrogen, USA) supplemented with 10% Fetal Bovine Serum (FBS) (Gibco) and at 37°C in a humidified 5% CO2 incubator. After subsequent expansion, MenSCs at passages 2-4 were characterized and cryopreserved for next experiments. Briefly, the test tubes containing 106 of the cultured cells in passages 2-4 were incubated individually with PE-conjugated monoclonal antibodies specific for CD29, CD45, CD73, CD105, and CD133 for 30 min at 4°C. Isotype-matched PE-conjugated antibodies were used as an isotype control. To evaluate OCT-4 expression, the cells were first fixed in 4% paraformaldehyde for 10 min and then washed. The cells were permeabilized with 0.1% saponin and washed and stained with rabbit anti-human OCT-4 antibody for 30 min, followed by 30 min incubation with FITC-goat anti-rabbit Ig (Avicenna Research Institute, Tehran, Iran). Considering the appropriate isotype controls, the cell populations were analyzed using a flow cytometer (Partec GmbH, Munster, Germany).
Isolation and cultivation of keratinocytes and fibroblasts: Isolation of keratinocytes and fibroblasts was performed according to protocols previously reported 27,28. Briefly, the foreskins were transferred in the tube containing Hanks' Balanced Salt Solution (HBSS) supplemented with 2.5 µg/ml amphotericin B (fungizone), 100 µg/ml streptomycin, and 100 U/ml penicillin (Sigma). Each sample was washed immediately with PBS and then minced. The samples were placed in the tube containing (0.25% v/v) dispase (Gibco) in HBSS and incubated at 4°C overnight. Next day, the contents of each individual tube were decanted into a petri dish while dermal side faced down. The epidermis and dermis were separated from each other using narrow-tipped forceps. The epidermal layers were chopped into small pieces and 2.5 ml trypsin/EDTA (0.25% trypsin, 1 mM EDTA, Gibco) was added. The dishes were incubated at 37°C in a humidified 5% CO2 incubator for 20 min, and finally to inactivate trypsin/EDTA, the dish contents were transferred into a tube containing 5 ml DMEM (Gibco) supplemented with 10% FBS and pipetted up and down. The suspension was passed through 70 µm mesh filter and then centrifuged at 200 g for 10 min. The resultant cell pellet was resuspended in Epilife medium (GIBCO Invitrogen, USA) and transferred into flasks coated by ECM protein. Primary keratinocytes were maintained at 37°C in a humidified 5% CO2 incubator and the medium was changed next day. In parallel, for fibroblast isolation, the dermal layers were minced into small pieces, and followed by digestion with 0.05% collagenase A (Roche Applied Science, Penzberg, Germany) in DMEM with 10 mg/ml human serum albumin (Sigma-Aldrich, MO, USA) during overnight at 37°C with 5% CO2. The suspension was then centrifuged at 400 g for 10 min and resuspended in Epilife medium. The primary fibroblasts were maintained at 37°C in a humidified 5% CO2 incubator and cultivated by replacing the culture medium twice weekly. Finally, isolated fibroblasts and keratinocytes were utilized for experiments before the sixth passage.
Fabrication of wound dressings: The bilayer scaffold composed of silk fibroin and Human Amniotic Membrane (HAM) was fabricated according to our established protocol (Figure 1A) 19. After fabrication, the circular bilayer scaffolds on the side of electro-spun silk fibroin were placed on round coverslips, put on the bottom of each well of 24-well cell culture plates and soaked with the complete culture medium for 30 min. After that, 150×103 MenSCs suspended in DMEM-F12 were transmitted. After keeping the plates in a humidified 5% CO2 incubator at 37°C for 45 min, 200 µl of complete DMEM-F12 medium was transferred into the wells for two turns, each after 1 hr interval. Then, the plates were preserved in CO2 incubator for overnight 20. The seeding of fibroblast/keratinocyte on bilayer scaffold was performed similarly to what is mentioned above with minor difference. 50×103 fibroblasts suspension in DMEM-F12 were dropped on the bilayer scaffolds and kept in a humidified 5% CO2 incubator at 37°C for 45 min. DMEM-F12 medium was added in two turns as alluded earlier and then the plates were incubated overnight. The next day after draining culture medium, 100×103 keratinocytes suspended in EpiLife medium were dropped on bilayer scaffolds and then the steps continued as described above (Figure 1B).
In vivo modelling: Forty-five male C57BL/6 mice were individually put in cages with controlled temperature (20±2°C) and humidity (50±10%). Pelleted diet (Behparvar Co, Iran) and water ad libitum were also administrated. Diabetes was induced according to our protocol 20 (Figure 1C). After confirmation of diabetes mellitus induction, animals were randomly allocated into three groups including: No-Treatment group (NT), fibroblast/keratinocyte-seeded bilayer scaffold treated group (bSC+FK), and MenSCs-seeded bilayer scaffold treated group (bSC+ MenSCs). Each group consisted of 15 mice and was equally divided into sub-groups according to the time points (3, 7, and 14 days after surgery).
Anesthesia was performed by intraperitoneal injection of 50 mg/kg ketamine (10% Ketamine, Alfasan Co., Woerden, Netherland) and 100 µg/kg medetomidine (Dorbene, syva, Spain). Full-thickness wounds were created by 5 mm biopsy punching (Kai Medical, Japan) on the back of mice seven days after induction of diabetes mellitus according to our protocol 20. After hemostasis, the wound dressings were applied on the wound sites (Figure 1D, step 2). Circle washer splints were set to wound edges using EPIGLU ethyl-2-cyanoacrylate tissue adhesive (Meyer-Haake, Germany), and then the splints were fixed to the skin using 5-0 nylon (Teb Keyhan Inc, Iran) sutures (Figure 1D, steps 3 and 4). 3 M Tegaderm® film (Health Care, Germany) dressing and cohesive bandage (Pet Flex cohesive bandage, Andover, Netherland) were applied around the wounds (Figure 1D, steps 5 and 6). Atipamezole (500 µg/kg) (Alzane, syva, Spain) was injected in order to facilitate the recovery phase. Tramadol (10 mg/kg) (Caspian Tamin Co, Iran) and Enrofloxacin (15 mg/kg) (Nasr pharma, Iran) were subcutaneously administered for three consecutive days’ post operation to increase the physiological recovery rate and relieve post-surgery pains.
Sample harvesting: At 3, 7, and 14 days post-surgery, complete wound area and approximately 2 mm of the margins of the wounds were punched 20. For histopathological studies, samples were collected in Neutral-Buffered Formalin (NBF) 10%, and for immunofluorescence staining, samples were snap-frozen in OCT (Killik, Bio-Optica, Milan, Italy). Then, embedded frozen blocks were stored at -80°C until cryo-section operation. The specimens for RNA isolation were immediately frozen in liquid nitrogen and then stored at -80°C.
Gross examination and calculation of the healing progression: Skin wound healing was evaluated by photographic images at the time of wound induction and at 3, 7, and 14 days’ post operation. Then, the Images were processed by ImageJ software (National Institutes of Health, Bethesda, MD). The percent of healing progression was calculated by following equation 29:
% Healing progression=[[(WA0) - (OWAn + EWAn)]/ (WA0)] ×100
(WA0): Wound area at the time of wound induction
(OWAn): Wound area at each time point consist of opening wound area
(EWAn): Epithelialized wound area at each time point
Histopathological evaluation: The Hematoxylin and Eosin (H&E) stained sections were evaluated by a veterinary anatomic pathologist. The dermal and epidermal heights of the healed and unwounded areas were also measured in H&E stained tissue sections at ×100 and ×400 magnifications, respectively.
Immunofluorescence staining assay: The diabetic wound samples were snap frozen and sectioned at a thickness of 7 µm and fixed in cold acetone, dry packed, and stored at -80°C. For evaluation of epidermal and dermal differentiation during wound healing, wound sections were incubated with anti-IVL, and anti-type III collagen, respectively as a primary rabbit polyclonal antibody, and then followed by an Alexa Fluor 488- donkey anti-rabbit IgG as a secondary antibody. The evaluation of angiogenic and neovasculogenic markers were performed by double immunofluorescence staining of the expression of CD31 and von Willebrand Factor (vWF) 30-34. The wound sections were initially incubated with, anti-vWF antibody and then continued by Alexa Fluor 488-conjugated donkey anti-rabbit IgG antibody. Following, the staining process was completed by PE-conjugated rat anti mouse CD31. Cutaneous innervation was visualized through intracellular immunolabeling of the pan-neuronal marker protein gene product 9.5 (PGP9.5) by rabbit anti mouse PGP9.5. The PGP9.5 labeled sections were incubated with secondary antibody, Alexa Fluor 488-conjugated donkey anti-rabbit IgG.
For numeration of neutrophils and macrophages in diabetic wound bed, double immunofluorescence staining was designed. Hence, sections were incubated in cocktail of two antibodies, PE-conjugated anti-mouse Ly-6G antibody and Alexa Fluor 488-conjugated anti-mouse F4/80 antibody, respectively. Macrophage characterization according to M1 or M2 polarization was performed through two sets of double immunofluorescence staining; the first evaluation by Alexa Fluor 488-conjugated anti-mouse F4/80 antibody and PE-conjugated anti-mouse CD38 antibody for M1 phenotype, and the second evaluation by Alexa Fluor 488-conjugated anti-mouse F4/80 antibody and PE-conjugated anti-mouse CD206 for M2 phenotype 35. In each evaluation, tissue nuclei were visualized with 4′6-diamidino-2-phenylindole (DAPI) (Sigma Chemical Company, St Louis, MO, USA). Five images were taken from the width of the wound at ×400 magnification after immunofluorescence staining. All images were individually analyzed using NIH ImageJ software as described in our previous report 20. The antibody profile utilized for immunofluorescence staining in this study is mentioned in table 1.
Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR): Isolation of total RNA was carried out from diabetic wound area and a thin margin surrounded it by using AccuZol TM total RNA extraction solution (Bioneer, Korea) according to the manufacturer’s instructions. Briefly, extracted total DNase-treated RNA (1 µg) was reverse transcribed into cDNA using random primers and thermocycler (Eppendorf, Germany). In next step, RT-qPCR analysis of gene expression was performed at Rotor-Gene Q System (Qiagen, CA) with the primers listed in table 2. All values were normalized against beta-2-micro-globulin as housekeeping genes and expressed as fold change or relative expression using the 2-ΔΔCT formula 36. Mean efficiencies and crossing point values for each gene were determined with LinReg PCR (version 11.0). The expression ratios of each bSC+MenSCs samples were calculated compared to other groups using relative expression software tool-2009 (REST-2009) (available at http://www.gene-quantification. info).
Statistical analysis: Results were presented as mean±standard error. Normally distribution of variables was evaluated by Kolmogorov-Smirnov normality test. Differences of means were analyzed by Two-way ANOVA, followed by Tukey’s post-test used for analysis of multiple comparison groups. Probability value p≤0.05 was considered statistically significant.
Results :
Identification of cell surface markers of MenSCs: Immunophenotypic analysis of surface markers revealed MenSCs are positive for CD29, CD73, and CD105 as MSCs markers and OCT-4 as an embryonic cell marker. However, they remain negative for hematopoietic stem cell markers such as CD45 and CD133.
Greater wound healing rate by application of bSC+MenSCs in comparison to bSC+FK: Gross evaluation showed that over time, an upward healing trend (of healing progress) was observed in all groups; however, this rate in bSC+MenSCs group was considerably greater than bSC+FK and NT groups at all-time points (Figures 2A and B).
Application of bSC+MenSCs improve histopathological alterations more effectively than bSC+FK: As shown in figure 3A, three days after wound induction, in the NT group, full-thickness skin defects remained approximately unchanged and the wound surface was covered by crust and necrotic debris. The newly formed dermis was thin and mainly composed of immature granulation tissue with infiltration of neutrophils with diffuse pattern. Application of bSC+FK exhibited that the epithelialization was initiated from edges of the wounds and scab yet existed on wound surface. Below the scabs, a thin and immature granulation tissue with mild to moderate inflammatory cells infiltration was seen. Few disorganized collagen bundles were observed in newly formed dermis. Application of bSC+MenSCs as wound dressing caused observable re-epithelization beneath the crusts. The thickness of granulation tissue was greater than those of the bSC+FK group and the neovascularization was observed in the papillary layer.
In NT group, seven days after wound induction, the defect was not filled completely, yet. The dermis was composed of immature granulation tissue and collagen fibrils arranged in a haphazard form in dermis. Application of fibroblast/keratinocyte-seeded on bilayer scaffold caused regenerated hyperplastic epidermis covering the wound surface. In sub epidermal layer, a highly vascularized and proliferating granulation tissue, consisting of more active fibroblasts, mild to moderate infiltration of inflammatory cells with slight superficial edema were detected. The connective tissue bundles were loosely arranged in the newly formed dermis. Using MenSCs-seeded bilayer scaffold initiated observable regeneration in both epidermal and dermal sites. The dermis was occupied with relatively mature granulation tissue and collagen bundles were arranged more regularly showing that dermal regeneration is boosted using MenSCs. Blood vessel density was noticeable in dermis. However, there were no signs of hair follicles or other adnexa formation in the regenerated dermis at this time point.
On day 14 after wound induction, cases belonging to the NT group demonstrated partial re-epithelization. Fibroblasts proliferation and mild to moderated infiltration of inflammatory cells in dermis were also seen. However, the wound sites were devoid of the skin adnexa. Histological examination of wounds in group applied by fibroblast/keratinocyte-seeded on bilayer scaffold 14 days after grafting revealed approximately well-recovered epidermal layers with more mature differentiation. The wound bed was mainly filled with fibroblast proliferation. Collagen bundles were arranged nearly parallel to the epidermis in the regenerated dermis. The inflammatory reaction was mild. Granulation tissue displayed reduction in the number of vessels mainly in the deeper layers of the wound. Histopathological evaluation showed that after application of MenSCs-seeded bilayer scaffold on wound site reasonably noticeable re-epithelization occurred. The new dermis was composed of relatively mature connective tissue and in some cases, well-recovered skin appendages were detected. The amount of inflammatory reaction was significantly scant (Figure 3A).
The mean Epidermal Thickness Index (ETI) was calculated at three time points for each study groups. The mean ETI of bSC+MenSCs group at day 3 was significantly higher than NT (p<0.05) and bSC+FK (p<0.05). Interestingly, a significant reduction of epidermal thickness to the optimal level was observed in bSC+MenSCs group from day 7 to 14 and at day 14, the epidermal thickness of this group was near to the thickness of adjacent epidermis. However, epidermal thickness in NT group was still measured at the lowest level (Figure 3B). The mean DTI of bSC+MenSCs group calculated at day 3 was significantly higher than NT (p<0.05) and bSC+FK (p<0.05), and at day 7 was also significantly higher than NT (p<0.0001) and bSC+FK (p<0.001). At the last time point (day 14), the measured mean DTI of bSC+MenSCs group showed that scar thickness considerably was restored to the optimal level (DTI £1). However, the dermal thickness in other groups was lower that native adjacent dermis (Figure 3C).
Identifying keratinocyte maturation in diabetic wounds dressed by the cell-seeded bilayer scaffolds: Normal differentiation during wound healing is characterized by restoration of expression of differentiation markers like Involucrin (IVL) in the neo-epidermis. Our data determined the higher expression of IVL by application of the cell-seeded bilayer scaffold in stimulation of diabetic wounds at day 7 and 14 in comparison to NT group. However, the IVL immunoreaction in bSC+MenSCs group was considerably higher than bSC+FK group and NT group at days 7 and 14 (Figures 4A and B).
Identifying type III collagen expression in diabetic wounds dressed by the cell-seeded bilayer scaffolds: In NT and bSC+FK groups, increasing expression of type III collagen was significantly determined from day 3 to day 14. However, in bSC+MenSCs group the expression pattern of collagen III demonstrated an upward trend from days 3 to 7 and then a downward trend from days 7 to 14 (Figures 5A and B).
Enumeration of inflammatory cells infiltration in diabetic wounds dressed by the cell-seeded bilayer scaffolds: In NT group, significant infiltration of neutrophils in wound site was noticed in all time points. However, conspicuous decline of neutrophilic infiltration in diabetic wound dressed by MenSCs-seeded bilayer scaffold occurred at day 14 and the reduction was more significant by application of bSC+MenSCs in comparison to NT group (p<0.001) and bSC+FK (p<0.05). The evaluation of macrophages showed that an increase in the count of macrophages in all groups occurred over time, but the total number of macrophages in treated groups was not remarkably different at each three time points. The total number of macrophages in NT group was significantly higher in comparison with diabetic wound dressed by MenSCs-seeded bilayer scaffold at day 14 (p<0.0001) (Figures 6A and B).
Furthermore, from day 3 to day 14, an increasing number of F4/80+CD38+ phenotype (M1 macrophage) was detected in NT and bSC+FK groups and at day 7, the mean cell count of F4/80+CD38+ phenotype in bSC+MenSCs group showed statistically significant expansion in comparison with NT group (p<0.01) and bSC+FK group (p<0.01). Remarkable reduction was determined in the mean cell count of F4/80+CD38+ phenotype in bSC+MenSCs group at day 14 compared to the NT and bSC+FK groups (p<0.05 and p<0.05, respectively) (Figures 7A and C).
While, there were no significant differences in the mean cell count of F4/80+CD206+ (M2 macrophage) phenotype among the different groups at day 3, the low frequency of F4/80+CD206+ phenotype was observed in NT and bSC+FK groups at day 7 and day 14. Interestingly, we determined a considerable increase in F4/80+CD206+ phenotype in the diabetic wound dressed by MenSCs-seeded bilayer scaffold comparing NT group and bSC+FK (p<0.0001 and p<0.0001 respectively) at day 7, and day 14 (Figures 7B and D).
The effect of cell-seeded bilayer scaffold wound dressings on neovasculogenesis and angiogenesis: CD31 expression pattern exhibited statistically significant increase in bSC+FK group in comparison to NT group at day 7 (p<0.0001) and day 14 (p<0.0001), respectively. However, the expression of CD31 and especially vWF in bSC+MenSCs group were considerably higher in comparison to other groups at three time points (Figures 8A-C).
The effect of cell-seeded bilayer scaffold wound dressings on innervation in diabetic wound healing: The PGP9.5 expression in diabetic wound sections was assessed through relative fluorescence intensity measurement of PGP9.5 immunofluorescence patterns. The PGP9.5 assessment of diabetic wound area represented statistically significant increase in bSC+ MenSCs group in comparison to other groups at day 7 and day 14 (Figures 9A and B).
Differential transcription pattern of wound healing related genes in different wound dressing: During wound healing in diabetic mice, the gene expression of some critical Inflammatory, immunomodulatory and angiogenic biomarkers were analyzed. IDO1 and Ptgs2 are implicated in immunomodulation and inflammation, respectively. There was also noteworthy increase in IDO1 expression ratio in bSC+MenSCs in comparison to NT and bSC+FK groups at three time points. On the other hand, bSC+MenSCs group displayed descending trend Ptgs2 expression in comparison to NT group and bSC+FK group at three time points. Egr2, the gene involved in M2 macrophage polarization showed statistically significant increase compared to NT group and bSC+FK group at day 3, day 7 and day 14.
Application of MenSCs-seeded bilayer scaffold on diabetic wound bed caused higher expression of CXCL12 in comparison to the NT group at three time points. Lower level of CXCL12 expression in bSC+MenSCs group in comparison to bSC+FK group was observed at days 3 and 7, However, significant expression occurred at day 14. Significant superiority of Mmp9 gene expression was identified in bSC+ MenSCs group in comparison to NT group at day 3 and bSC+FK group at all-time points. Furthermore, up-regulation of Vascular Endothelial Growth Factor A (VEGFA) mRNA level was sharper in MenSCs-seeded bilayer scaffold compared to NT group, and bSC+FK group at three time points (Figure 10).
Discussion :
In the present study for the first-time, we applied bilayer scaffold composed of AM and silk fibroin loaded with fibroblast/keratinocyte for healing of the diabetic wound in comparison to what is known as enhanced regenerative properties of MenSCs integrated in bilayer scaffold. There are several reports that indicate that keratinocyte and fibroblast interactions have central role in the wound healing and fibroblast differentiation potentially affects epithelial wound closure and scar remodeling 9. However, in impaired diabetic wound, healing progression was significantly higher when MenSCs-seeded bilayer scaffold wound dressing was used in comparison to the fibroblast/keratinocyte-seeded bilayer wound dressing. Pertaining to epithelialization as an essential phase of the wound healing process, expression of IVL was assessed as a marker of migrating differentiated keratinocytes. As mentioned before, IVL expression was determined significantly higher in bSC+FK group in comparison to NT group at day 7 and day 14 and probably related to the reported properties of keratinocytes-seeded on bilayer scaffold that integrated into wound bed 37. Interestingly, IVL expression was considerably higher in MenSCs seeded bilayer scaffold wound dressing in comparison with other groups at day 7 and day 14. In previous studies 38,39, we indicated that MenSCs could potentially differentiate into mature keratinocytes and express several epidermal specific markers such as K14, p63, and IVL. Collectively, it can be concluded that higher expression level of IVL in bSC+MenSCs group might be also considered as trans-differentiation of MenSC phenotype into keratinocytes in diabetic wound bed dressed by MenSCs-seeded bilayer scaffold. Moreover, dermal thickness in bSC+MenSCs group represented noteworthy increase during 7 days after wound induction in comparison to bSC+FK and NT groups. H&E examination confirmed increased DTI index in bSC+MenSCs group and decreasing dermal cellularity to normal level same as adjacent skin at day 14. However, dermal thickness in diabetic wound site in NT and bSC+FK groups were less than normal adjacent skin after 14 days, thus low proliferation rate and ECM deposition contribute to impaired wound healing. These resultsconfirm that sustained inflammation in chronic wounds, lead to fibroblast dysfunction 40. Hence, in line with our findings, seeded fibroblast/keratinocyte on bilayer scaffold wound dressing has not been as effective as MenSCs-seeded bilayer scaffold in progression of regenerative procedures in diabetic wounds.
In early phase of wound healing, deposition of ECM proteins particularly type III collagen happens in dermis 41,42 and consistent with our previous findings 20. In early phase of wound healing, deposition of ECM proteins particularly type III collagen happens in dermis 41,42. In consistent with our previous findings, contrary to upward trend of total collagen from day 3 to day 14 in other groups, MenSCs-seeded bilayer scaffold wound dressing inversely showed downward trend of type III from day 7 to day 14 20. Therefore, accumulation of type I collagen deposition has been proposed as a characteristic of newly formed dermis maturation 43. Our data showed that maximum dermal hypertrophy and expression of type III collagen were observed in bSC+MenSCs group at day 7 and further at day 14 more than two-fold reduction was determined in type III collagen density. In line with these findings, it was reported that recruitment of MenSCs in normal wound healing in mice lead to deposition of well-organized collagen fibers in dense thick bundles following reduction of type III collagen proportion 31, as well as MMPs during remodeling phase. MMP-9 play a detrimental role in diabetic wound healing 44 and the evaluation of MMP-9 expression can be an accurate prediction of the success in treatment of the chronic wound 45.
Consistent with previous data, our results exemplified by diabetic wound dressed by MenSCs seeded bilayer scaffold, the level of MMP-9 mRNA expression was significantly decreased from day 7 to day 14 in comparison to NT group and FK+bSC group. However, we observed that lower level of the MMP-9 mRNA along with fibroblast/keratinocyte-seeded bilayer scaffold wound dressing in comparison to no treated wounds at day 7 after diabetic wound induction, is likely to be related to the short-term role of fibroblasts implanted on bilayer scaffold in modulating inflammation through the secretion of protease inhibitors, including TIMP-1 and -2 46-48.
In diabetic wound, it has been reported that inflamed wound chronicity and oxidative stress lead to premature senescence and irreversible arrest of proliferation in fibroblasts 49. Furthermore, keratinocytes in the chronic wound are unable to upregulate chemokines and cytokines are necessary for the resolution of inflammation 50 and in diabetic wound due to prolonged inflammation phase, excessive infiltration of leukocytes lead to non-healing wound process 51-53. Our data designated that the recruitment of fibroblast/ keratinocyte-seeded bilayer scaffold could considerably reduce inflammation at day 3 and day 7 in comparison to NT group, but at day 14 according to prolonged inflammatory state and sustained inflammatory phase there were no significant differences in the amount of the neutrophils between bSC+FK and NT groups. On the other hand, the diabetic wound dressed by MenSCs-seeded bilayer scaffold showed a significantly downward trend of neutrophil accumulation in wound bed in comparison to NT group in all studied time points. Furthermore, bSC+MenSCs group showed the lowest numbers of neutrophils in comparison with bSC+FK and NT groups at day 14. Collectively, its seems that due to non-resolving chronic inflammation, keratinocytes and fibroblasts represented impaired ability to upregulate anti-inflammatory cytokines and chemokines and this cell-based bioengineered wound substitute could not promote delayed wound healing 40,50.
In consistent with previous results, noticeable distribution of M1 macrophages was detected in the diabetic wound site in bSC+FK and NT groups during 14 days. However, the lowest number of M1 phenotype was noticed in diabetic wound dressed by MenSCs-seeded bilayer scaffold at day 14. Conversely, the greatest number of M2 phenotype presented in wounds treated with MenSCs-seeded bilayer scaffold at day 7 and day 14. In line with these findings, we determined significantly higher Egr2 mRNA expression (the gene involved in M2 macrophage polarization) at day 7 and day 14 in bSC+MenSCs group in comparison to other groups. According to previous reports 40,54, prolonged inflammation in the chronic wound bed might lead to fibroblast dysfunction.
Furthermore, Galkowska et al found that keratinocytes and endothelial cells in the chronic wound site are unable to upregulate necessary chemokines and cytokines, and that in turn, continual elevation of Reactive Oxygen Species (ROS), might also occur 50. Overproduction of ROS lead to ECM and cell membrane damage 55, activation of proteases including MMPs and serine proteases and inactivation of protease inhibitors that result in premature cell senescence 56-58. In fact, we represented that fibroblast/keratinocyte-seeded bilayer scaffold wound dressing failed to resolve inflammation and wound resolution.
In line with these findings, we also analyzed differential Ptgs2 mRNA expression among study groups, while significantly ascending trend of cyclooxygenase (COX)-2 expression (Ptgs2) was determined during 14 days in no treatment group and diabetic wound dressed by fibroblast/keratinocyte-seeded bilayer scaffold. In contrast, significantly descending trend of Ptgst2 in diabetic wound dressed by MenSCs-seeded bilayer scaffold indicated a substantial immunomodulatory effect of MenSCs in combination with bilayer scaffold applied in the wound site. We also evaluated the IDO1 mRNA expression to identify the immunomodulatory role of MenSCs-seeded bilayer scaffold recruited in diabetic wound 59,60. We observed that IDO1 expression in diabetic wound bed significantly up-regulated in bSC+MenSCs group especially in comparison to bSC+FK group and NT group during 14 days after diabetic wound induction.
In diabetic wound healing, successful wound revascularization failed to progress 61. The results exhibited that noteworthy increased expression of CD31 occurs in bSC+MenSCs group as well as in bSC+FK group in comparison to no treatment group at day 7 and day 14. Although, diabetic wounds dressed by MenSCs-seeded bilayer scaffold presented higher expression of CD31 even in comparison to bSC+FK group from day 7 to day 14. Interestingly, the strongest expression of vWF in diabetic wounds were identified when wounds were dressed by MenSCs-seeded bilayer scaffold at three time points. We also assessed CXCL12 or SDF-1 mRNA expression level in diabetic mice wounds as a principal chemokine involved in recruiting of Endothelial Precursor Cells (EPCs) into the wound site 62. In agreement with immunofluorescence evaluation of CD31, the qRT-PCR analysis also showed significant expression of CXCL12 mRNA in bSC+MenSCs group in comparison with no treated diabetic wounds. Hence, the immunomodulatory potency and specific paracrine effect of MenSCs in combination with bilayer scaffold properties resulted in reducing prolonged inflammation of diabetic wound and enhance angiogenesis and neovascularization 31,32,63 or even their differentiation into mature endothelial-like cells 33,34.
To our knowledge, polarization of macrophages from pro-inflammatory M1 phenotype into anti-inflammatory M2 phenotypes occur in the latter stages of wound healing 35,64. M2 macrophages that represent pro-healing functions, play a critical role in epithelialization and innervation in wound healing through producing anti-inflammatory and pro-resolving neuroprotectin/protectin D1 (NPD1/PD1) 65. Persistent chronic inflammation in diabetic wounds causes excessive or unresolved M1 macrophage activation which contributes to the impairment of wound healing and repairing of injured peripheral nerves, in turn results in diabetic peripheral neuropathy and pain insensitivity 66,67. In this study, the up-regulation of PGP9.5 was significantly observed in diabetic wound dressed by MenSCs-seeded bilayer scaffold in comparison to other groups at day 7 and day 14. Consistent with this data, it has been shown that macrophages produce anti-inflammatory and pro-resolving NPD1 involved in attenuating macrophage inflammatory activities, chronic inflammation and oxidative stress after unresolved inflammation in diabetic wound 65.
Collectively, our results indicate that MenSCs in conjunction with bilayer scaffold provide a specialized stem cell‐derived skin substitute and exhibits a complex of immunomodulatory, clonogenicity, migratory and specific paracrine signature necessary to enhance regeneration in non-healing diabetic wound environment. Fibroblast/keratinocyte-seeded bilayer scaffold wound dressing was considered as a compensatory reparative population, however in diabetic wounds, collagen remodeling and ECM formation are prevented and fibroblasts and keratinocytes exhibit reduced reaction to growth factors, lower migratory ability, and increased apoptosis rate 40,68. Also, inability of fibroblast and keratinocyte to express anti-inflammatory functions because of persistence of inflammatory state in diabetic wound microenvironment resulted in irreversible growth arrest or replicative senescence in quiescent fibroblasts settled in wound bed 40. Consequently, senescent fibroblast probably could not contribute to mobilization of EPCs from bone marrow niches toward the wound site and homing to diabetic wound bed 69. In fact, chronic inflammation 70,71 and bacterial infection 72,73 play contributory roles in scar formation and insufficient regeneration in diabetic wound bed and fail to obtain satisfactory results 74,75.
Conclusion :
This study shows that MenSCs with their superior biological properties, including potent immunomodulatory, proangiogenic and reparative potential in combination with bioengineered bilayer scaffold skin substitute not only enhanced diabetic wound healing and cutaneous regeneration process in vivo, but also may be relevant for the clinical application and treatment of diabetic impaired wound healing process. From the current study, it seems that fibroblast and keratinocyte cells-loaded bilayer scaffold like commercial cell-based skin substitutes are unable to attenuate pro-inflammatory microenvironment in diabetic mice model and promote impaired wound healing process. Future research should be conducted to explore the specific mechanisms involved in the enhanced immunomodulatory and reparative properties of MenSCs on chronic wound healing process.
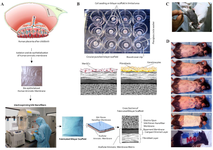
Figure 1. Steps of preparation and application of wound dressing in diabetic mice. A) Human placenta being surrounded by two layers of the amniotic membrane and chorion. The human amniotic membrane was separated from amniochorionic membrane, de-epithelialized and freeze-dried. Nano-fibers were electro-spun on the basement membrane side of de-epithelized HAM by pumping silk fibroin solution by high voltage transmission. The schematic cross section of bilayer scaffold was showed. B) Cultivation of MenSCs and/or fibroblasts/keratinocytes for seeding on bilayer scaffold through sequential steps as mentioned in earlier report. C) Induction of diabetes by streptozotocin in C57BL/6 mice, D) Different parts of intervention (steps 1 to 6). 1) induction of full-thickness skin wounds, 2) wound dressing implantation, 3 and 4) splinting toward 5 and 6) wound dressing and bandage.
|
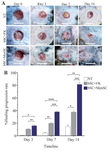
Figure 2. Gross evaluation of wound healing progression. A) Macro photographs of wounds from one selected mouse in each group in three time points. B) The percent of wound healing progression during three time points (p-values less than 0.05, 0.01, 0.001, and 0.0001 were shown with *, **, ***, and ****, respectively.
|
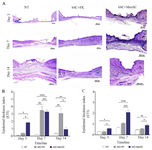
Figure 3. Microscopical evaluation of wound healing. A) H & E staining of healed wounds in different groups. Day 3: In NT group, lack of epidermal regeneration (black arrow), scab formation (red arrow) and thin granulation tissue in dermis (black star), are noted. In bSC+FK group, denuded wound covered by fibrino necrotic exudate (black arrow) in the central part and inflammatory cell infiltrations (black star) in newly-formed dermis is noticeable. In bSC+MenSCs group, sign of regeneration (white arrow) specially in wound borders beneath the remnant of wound dressing (black arrow) is detected. Also note the immature and edematous granulation tissue (green star) in dermis. Day 7: in NT group, the epidermal regeneration is not complete (black arrow) and also note to hyper cellular granulation tissue in dermis mainly occupied with inflammatory cells (black star). In bSC+FK group, note to regenerated hyperplastic and stratified epidermis (black arrow) and proliferating granulation tissue (black star). In bSC+MenSCs group, the re-epithelization is almost acceptable (black arrow) and in dermis a hyper cellular and edematous granulation tissue (black star) with noticeable vascular density is observed. Day 14: in NT group the newly formed epidermis was characterized by immature keratinocytes (black arrow) and new dermis was considered by an immature granulation tissue (black star). In bSC+FK group, the re-epithelization is not complete specially in central part (black arrow) and in dermi, note to hyper cellular granulation tissue (green arrow). In bSC+ MenSCs group, more mature stratified squamous epithelium (black arrow), dense collagenous bundles (green star) and well-recovered skin appendages (green arrow) are noted. B and C) The averaged Dermal Thickness Index (DTI) and Epidermal Thickness Index (ETI), respectively. p-values less than 0.05, 0.01, 0.001, and 0.0001 were shown with *, **, ***, and ****, respectively.
|
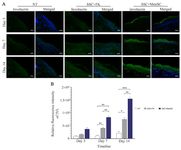
Figure 4. Evaluation of IVL expressions in the study groups. A) IVL expression in diabetic wounds was visualized by immunofluorescence staining through Alexa Fluor®488 labeling of IVL. These sections were also counterstained with DAPI. Scale bar = 50 µm. B) Relative fluorescence intensity of IVL expression of diabetic wounds was compared in different groups. p-values less than 0.05, 0.01, 0.001, and 0.0001 were shown with *, **, ***, and ****, respectively.
|
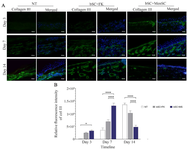
Figure 5. Evaluation of Type III collagen expressions in the study groups. A) Type III collagen levels in diabetic wounds was visualized by immunofluorescence staining through Alexa Fluor®488 labeling of Type III collagen. These sections were also counterstained with DAPI. Scale bar = 50 µm. B) Relative fluorescence intensity of type III collagen expression of diabetic wound. p-values less than 0.05, 0.01, 0.001, and 0.0001 were shown with *, **, ***, and ****, respectively.
|
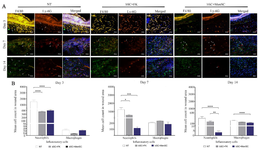
Figure 6. Neutrophil and macrophage enumerations in diabetic wound area. A) Neutrophil and macrophage were labeled by double immunofluorescence staining of diabetic wound sections by PE conjugated anti-Ly-6G/Ly-6C (Gr-1) targeted neutrophils (white arrows) and Alexa Fluor®488 conjugated anti-F4/80 targeted macrophages (white arrowhead). These sections were also counterstained with DAPI. Scale bar = 50 µm. B) Quantitative analysis of inflammatory cell enumerations at three time points among all groups. p-values less than 0.05, 0.01, 0.001, and 0.0001 were shown with *, **, ***, and ****, respectively.
|
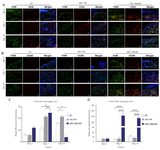
Figure 7. Effect of diabetic wound dressing on M1 and M2 macrophage subpopulations. A) M1 macrophages were labeled by double immunofluorescence staining of diabetic wound sections for Alexa Fluor®488 conjugated anti-F4/80 and PE conjugated anti-CD38. B) M2 macrophages were labeled by double immunofluorescence staining of diabetic wound sections through Alexa Fluor®488 conjugated anti-F4/80 and PE conjugated anti-CD206. Nuclei were visualized with DAPI (scale bar = 50 µm). C) Comparative evaluation of M1 phenotype subpopulation in diabetic wound treated in study groups. D) Comparative evaluation of M2 phenotype subpopulation in diabetic wound treated in study groups. p-values less than 0.05, 0.01, 0.001, and 0.0001 were shown with *, **, ***, and ****, respectively.
|
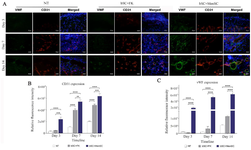
Figure 8. Angiogenesis and Vasculogenesis evaluations in diabetic wound healing. A) Diabetic wound sections were labeled by double immunofluorescence staining of CD31 and vWF expression pattern through PE conjugated anti-CD31 and Alexa Fluor®488 conjugated anti-vWF. These sections were also counterstained with DAPI. Scale bar = 50 µm. Comparative evaluation of relative fluorescence intensity of CD31 expression, B) and vWF expression, C) were evaluated on days 3, 7 and 14 in diabetic wounds. p-values less than 0.05, 0.01, 0.001, and 0.0001 were shown with *, **, ***, and ****, respectively.
|
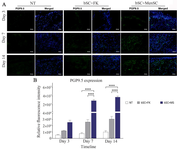
Figure 9. Evaluation of innervation pattern in diabetic wound healing. A) Diabetic wound sections were labeled by immunofluorescence staining of PGP9.5 expression pattern through Alexa Fluor®488 conjugated anti- PGP9.5. Indirect immunofluorescence staining for innervation assessment targeted diabetic wounds. These sections were also counterstained with DAPI. Scale bar = 50 µm. B) Comparative evaluation of relative fluorescence intensity of PGP9.5 expressions in diabetic wound. p-values less than 0.05, 0.01, 0.001, and 0.0001 were shown with *, **, ***, and ****, respectively.
|

Figure 10. Relative gene expression ratio of some wound healing related genes by different wound dressing. Matrix early growth response 2 (Egr2), metallopeptidase 9 (Mmp9), chemokine (C-X-C motif) ligand 12 (CXCL12 or SDF-1), indoleamine 2,3-dioxygenase 1 (IDO1), prostaglandin-endoperoxide synthase 2 (Ptgs2), vascular endothelial growth factor A (VEGFA) in diabetic wound areas of mice. The expression is normalized to the reference gene (B2m). There were five cases in each group. p-values less than 0.05, 0.01, 0.001, and 0.0001 were shown with *, **, ***, and ****, respectively.
|
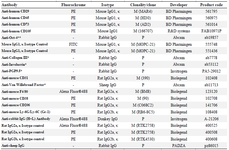
Table 1. Antibodies
* This antibody has specific reactivity with mouse and some other species. P: Polyclonal; M: Monoclonal.
** This antibody has specific reactivity with Mouse, Sheep, Human, Rhesus monkey.
|
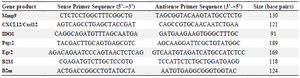
Table 2. Sequence of primers used in qRT-PCR
Mmp9: Matrix metallopeptidase 9; CXCL12: Chemokine (C-X-C motif) ligand 12; IDO1: Indoleamine 2,3-dioxygenase 1; Ptgs2: Prostaglandin-endoperoxide synthase 2; Egr2: Early growth response 2; B2m: Beta-2-microglobulin.
|
|