Investigating the Effects of HMGB1 Overexpression on Colorectal Cancer Cell Migration via Oncolytic Herpes simplex Virus Type 1 (oHSV-1)
-
Shayan, Sara
-
Department of Molecular Virology, Pasteur Institute of Iran, Tehran, Iran
-
Arashkia, Arash
-
Department of Molecular Virology, Pasteur Institute of Iran, Tehran, Iran
-
Bahramali, Golnaz
-
Department of Hepatitis and AIDS and Blood Borne Diseases, Pasteur Institute of Iran, Tehran, Iran
-
 Azadmanesh, Kayhan
Department of Molecular Virology, Pasteur Institute of Iran, Tehran, Iran , Tel: +98 21 66953311-20; E-mail: azadmanesh@pasteur.ac.ir
Azadmanesh, Kayhan
Department of Molecular Virology, Pasteur Institute of Iran, Tehran, Iran , Tel: +98 21 66953311-20; E-mail: azadmanesh@pasteur.ac.ir
Abstract: Background: Colorectal Cancer (CRC) represents a significant global health challenge, and its progression, resistance to therapy, and metastasis are strongly influenced by the tumor microenvironment, including factors like hypoxia. This study explores the impact of High Mobility Group Box 1 (HMGB1) overexpression on CRC cell migration, while identifying potential genes associated with this process.
Methods: To explore this, we developed oncolytic virotherapy, resulting in HSV-HMGB1, an oncolytic Herpes simplex virus that expresses HMGB1. HMGB1 is known its role in cancer progression, particularly in the context of cancer cell migration.
Results: Contrary to expectations, our scratch assays indicated that HSV-HMGB1 did not significantly induce migration in CRC cells, suggesting that HMGB1 might not directly contribute to this process. Employing microarray analysis, we investigated gene expression changes linked to CRC cell migration, leading to construction of a Protein-Protein Interaction (PPI) network. This network revealed the presence of hub proteins, including as NDRG1, LGALS1, and ANGPTL4, which are recognized for their roles in cancer cell migration. The differential expression of these genes under hypoxic conditions was further validated using quantitative RT-PCR, aligning with the findings from our microarray data.
Conclusion: Our findings emphasize the complex regulation of CRC cell migration, and provides valuable insights into potential molecular mechanisms and pathways. These findings have implications for further research into cancer progression and the development of therapeutic strategies.
Introduction :
Colorectal Cancer (CRC) is a major health issue and ranks among the primary causes of cancer-associated mortality globally 1,2. CRC is the third most common cancer worldwide, accounting for approximately 10% of all cancer cases and is the second leading cause of cancer-related deaths worldwide 2. The tumor microenvironment, with its key components such as hypoxia, plays an essential role in cancer progression, resistance to therapy, and the development of metastasis 3,4. Hypoxia is a common characteristic of solid tumors, including CRC, and is caused by inadequate oxygen supply to the tumor cells due to their uncontrolled growth and poor vascularization 5,6.
Hypoxia has been closely associated with the metastatic potential of cancer cells. Studies have shown that the low-oxygen microenvironment not only fosters cancer cell survival and proliferation, but also enhances their ability to migrate and establish secondary tumors in distant organs 7. This multifaceted role of hypoxia in cancer progression underscores the need for innovative therapeutic strategies, such as oncolytic virotherapy, to effectively combat metastatic CRC.
Oncolytic virotherapy is an emerging treatment approach that uses viruses to selectively target and kill cancer cells 8. However, hypoxia poses a significant challenge to the effectiveness of oncolytic virotherapy because viral replication and spread are hindered in low oxygen environments 9,10. To overcome this barrier, researchers have explored different strategies, such as using hypoxia-activated prodrugs or modifying viruses to enhance their replication and spread under hypoxic conditions 11,12.
In our previous study, we developed an oncolytic Herpes simplex virus type 1 named HSV-HMGB1, which expresses the High Mobility Group Box 1 (HMGB1) protein 13. HMGB1 is a multifunctional protein involved in DNA repair, transcriptional regulation, and immune response 14,15. We previously demonstrated that HSV-HMGB1 showed more cytotoxicity in CRC cells under normoxic and hypoxic conditions than a control virus named HSV-ble in vitro 13.
HMGB1 is a protein with multifaceted functions, including its involvement in DNA repair, transcriptional regulation, and immune responses. Recent research has also implicated HMGB1 in cancer progression, particularly in the context of promoting cancer cell migration. Several studies have reported that HMGB1 can act as a potent inducer of cancer cell migration, facilitating the dissemination of cancer cells from primary tumor sites to distant locations within the body 16-18. This pro-migratory effect of HMGB1 has raised significant interest in understanding its role in cancer metastasis and its potential implications for therapeutic interventions. Although previous study has suggested that expression of HMGB1 might promote cancer cell migration and metastasis, our objective was to investigate whether overexpression of HMGB1 via HSV-HMGB1 could induce migration in CRC cells.
In conclusion, both hypoxia and the presence of HMGB1 protein have been linked to the induction of cancer cell migration. Given the clinical relevance of this issue, we embarked on a comprehensive investigation to determine whether the exposing CRC cells to hypoxic conditions or overexpressing HMGB1 via the oncolytic Herpes simplex virus led to enhanced cancer cell migration.
Materials and Methods :
Cell culture: CRC cell lines HT29, HCT116, and SW480 sourced from the National Cell Bank of Iran (Pasteur Institute of Iran), were cultured in Dulbecco's Modified Eagle Medium (DMEM) supplemented with 10% Fetal Bovine Serum (FBS) and 1% penicillin-streptomycin. Cells were maintained in a humidified atmosphere containing 5% CO2 at 37°C. To adapt the cells to hypoxic conditions, they were placed in an Anoxomat chamber (Mart Microbiology, Lichtenvoorde, The Netherland) (1% O2) for 4 hr and then they were transferred to a standard culture environment (5% CO2, 95% air) at 37°C for 48-72 hr. These adaptations were repeated twice weekly, and after 20 hypoxic exposures, the cell lines became hypoxia-conditioned.
Generation of HSV‐HMGB1 and HSV-ble: The oncolytic Herpes simplex virus type 1 (HSV) expressing the HMGB1 protein, denoted as HSV-HMGB1, and the control virus referred as HSV-ble, were constructed in accordance with the methods outlined in our prior publication 13. Briefly, the process involved cloning the HMGB1 gene into the into the HSV-1 backbone using homologous recombination. The control virus was generated using a similar approach, with the exception that it did not involve the insertion of HMGB1.
Cell migration assay: Cell migration was evaluated through a scratch assay. To initiate the assay, HT29, HCT116, and SW480 cells were seeded at 4.0×104 in 96-well plates and allowed to grow into confluent monolayers. Subsequently, a sterile pipette tip was used to create consistent scratches across these monolayers, thereby simulating a wound-like scenario. Once the scratches were established, the cells were infected with either HSV-HMGB1 or HSV-ble at MOI 1, under hypoxic and normoxic conditions. Images of the scratched areas were captured at 0-hr mark and again at the 24-hr juncture, using an inverted microscope to observe the migration of cells. Over the designated time period, the width of the scratched area was measured to quantitatively assess the degree of cell migration in response to HMGB1 overexpression. The analysis of these images was performed through the utilization of Image J software, and the percentage of wound confluence was determined through the following formula:
Wound confluence (%)=Area of original wound-area of wound during healingArea of original wound ×100
Microarray data and identification of differentially expressed genes (DEGs): GSE9234, a microarray expression profile dataset submitted by Guimbellot JS et al 19, and GSE124118, another microarray expression profile dataset, were obtained from the GEO database (http://www.ncbi. nlm.nih.gov/geo). The GSE9234 dataset included three samples of hypoxic- conditioned HT29 cells and three samples of normoxic-conditioned HT29 cells. This dataset was based on the GPL570 platform, utilizing the Affymetrix Human Genome U133 Plus 2.0 Array. The GSE124118 dataset contained three samples of MRC5 cells and three samples of MRC5 infected with HSV-1, built on built on the GLP18573 Illumina NextSeq 500. For analysis in this study, the microarray data was first normalized and back- ground-corrected. Subsequently, limma package, implemented in the R programming, was used to identify Differentially Expressed Genes (DEGs) between the hypoxic cells and their oxygenated counterparts 20 (Supplementary 1). Benjamin & Hochberg method was used to adjust p-values, which were calculated separately for each DEGs 21. A significant threshold of FDR<0.05 and a log Fold Change (FC) greater than 1.5 were used as the criteria for identifying DEGs.
Construction of a PPI network and analysis: In this study, Search Tool for the Retrieval of Interacting Genes (STRING https://string-db.org/cgi), an online database that predicts protein–protein interactions, was used to construct a PPI network of identified DEGs. A confidence score of 0.4 was set as the cut-off criterion 22. The confidence score of 0.4 was selected as the threshold for constructing the Protein-Protein Interaction (PPI) network in our study with the specific aim of identifying any potential nodes associated with cancer cell migration. The protein interaction network was visualized using NetworkAnalyst (https://www.net-workanalyst.ca) and Cytoscape (3.9.1) 23. To evaluate the nodes in the PPI network, we adopted several topological measures, including degree (k), Maximal Clique Centrality (MCC), and Betweenness Centrality (BC). Since degree (k), BC, and MCC are often used for detecting the hub in a network 24-26, we determined hub genes based on connectivity degree (number of interactions), MCC, and BC using Cytohubba on Cytoscape 27.
Functional and pathway enrichment analyses: Gene Ontology database (GO; www.geneontology. org) analysis 28, and Kyoto encyclopedia of Genes and Genomes (KEGG) 29 pathway enrichment analyses were performed using DAVID. Database for Annotation, Visualization and Integrated Discovery (DAVID) 6.8 (http://david.abcc.ncifcrf.gov/) is an online analysis platform that provides comprehensive biological and functional information associated with an extensive gene list 23. p<0.05 was considered to indicate a statistically significant difference.
RNA isolation and qRT-PCR: Trizol reagent (TaKara, Kusatsu, Shiga, Japan) was used for RNA isolation from the cells during normoxia and hypoxia. RNA samples were reversely transcribed to complementary DNA by the QIAGEN Reverse Transcription Kit (Qiagen, Germany). Subsequently, the quantification of cDNA was performed by the qRT- PCR method using SYBR Green Master Mix (Yektatajhiz, Iran). The reaction conditions were conducted at 95°C for 10 min, 40 cycles of 95°C for 10 s, and 60°C for 30 s. The RPLP0 was used as an internal reference control 30. Gene expression levels were calculated based on the Delta-Delta Ct relative quantification.
Statistical analysis: Statistical analyses were performed using the student’s t-test with GraphPad Prism 8 software (GraphPad Prism, San Diego, CA, USA). The p value was considered statistically significant when it was less than 0.05.
Results :
Effect of HSV-HMGB1 on CRC cell migration: To assess the impact of HMGB1 overexpression on CRC cell migration, we conducted scratch assays on HT29, HCT116, and SW480 cells under both normoxic and hypoxic conditions. Cells were infected with HSV-HMGB1, which resulted in HMGB1 overexpression, or the control virus HSV-ble as a negative control. After the scratch was made, the gap filling was assessed at 24 hr post-scratch. Surprisingly, we observed no significant migration in HSV-HMGB1-infected CRC cells under any tested conditions, including hypoxic environments. In contrast, uninfected CRC cells demonstrated migration under hypoxic conditions, suggesting that hypoxia alone can induce migration in all three cell lines (Figure 1).
Additionally, no migration was detected in the control cells infected with a control virus (HSV-ble) under both normoxic and hypoxic conditions. Our results indicate that HMGB1 overexpression in HSV-infected CRC cells did not induce migration, suggesting that HMGB1 alone may not play a direct role in promoting cell migration in CRC.
Protein–protein interaction (PPI) network analysis and pathway enrichment: Using the STRING database, a PPI network obtained from 3433 DEGs, which was composed of 296 nodes and 316 edges, was constructed and visualized in Cytoscape and NetworkAnalsyt (Figure 2A). Notably, within this network, we identified nine hub proteins, namely UBC, NDRG1, ENO2, LGALS1, NDUFA4L2, ANGPTL4, WRNIP1, ALDOC, and PCK1 based on various criteria such as degrees, MCC, and BC (Figure 3). Notably, we also generated a PPI subnetwork specifically focusing on the centrality of HMGB1 (Figure 2B). This subnetwork revealed that HMGB1 did not exhibit interactions with any of the identified hub proteins, suggesting a distinct role for HMGB1 within this network.
Gene Ontology (GO) analysis was performed using DAVID (Figure 4A). GO covers the three categories, including Cellular Component (CC), Biological Process (BP), and Molecular Function (MF). The nine hub genes were significantly enriched in gluconeogenesis, glycolytic process and cellular response to hypoxia (ontology: BP), extracellular exosome and extracellular space (ontology: CC), and magnesium ion binding (ontology: MF). Moreover, based on KEGG pathway analysis 6 pathways were significantly enriched among the hub genes (Figure 4B).
Validation of differentially expressed genes by quantitative RT‐PCR (qRT‐PCR) analysis: To investigates the role of migration-related genes among the hub genes identified through GO and KEGG pathway analysis, we focused on NDRG1, LGALS1, and ANGPTL4 for further analysis via qRT-PCR. The aim of this analysis was to determine whether the mRNA expression levels of these genes in HT29 cells infected with HSV-HMGB1 and HSV-ble were altered under both hypoxic and normoxic conditions. The results revealed that NDRG1, LGALS1, and ANGPTL4 were differentially expressed under hypoxic conditions and were consistent with microarray dataset results. NDRG1, LGALS1, and ANGPTL4 mRNA levels upregulated in HT29 cells under hypoxic conditions, but expression levels of these genes remained unchanged in HSV-HMGB1 and HSV-ble infected cells under normoxic and hypoxic conditions (Figure 5).
Integration of additional datasets to enhance hub gene analysis: The overall design of this section is illustrated in figure 6. In pursuit of a comprehensive understanding of hub gene dynamics in cancer cells infected with HSV-HMGB1 cell migration, we extended our analysis by incorporating additional datasets (GSE124118). We initially identified genes associated with migration through the NCBI database and cross-referenced these findings with our existing microarray dataset (GSE9234). Subsequently, we explored a distinctive context by examining a dataset comprising MRC5 cells (human diploid cell line), encompassing both control and Herpes simplex virus (HSV)-infected cells. This integration facilitated a more detailed exploration of hub gene behavior, with MRC5 data being chosen due to its availability as the sole dataset capturing the influence of herpes infection on cancer cells. Collectively, the amalgamation of these diverse datasets offers a more comprehensive perspective on the intricate mechanisms governing CRC cell migration. In the MRC5 dataset (GSE124118), our analysis revealed that NDRG1 and LGALS1 maintained their stability in infected cells, confirming our real-time PCR results. Notably, ANGPTL4 exhibited a modest reduction in infected MRC5 cells, in contrast to the upregulation observed under hypoxic conditions. This dual-context analysis provides insights into the complex gene regulation dynamics induced by HSV-1 (Figure 6).
Discussion :
In this study, we aimed to investigate the impact of HMGB1 overexpression using HSV-1 on CRC cell migration and to identify potential migration-related genes through PPI network analysis and pathway enrichment under hypoxic conditions. CRC cells were infected with the virus at a Multiplicity of Infection (MOI) of 1 for a duration of 24 hr. We conducted HSV-HMGB1 infection for 24 hr because this duration was sufficient for achieving overexpression of HMGB1 13. Moreover, extending the infection beyond 24 hr, such as at 48 and 72 hr, resulted in significant cell death in the CRC cells. This phenomenon can be attributed to several factors including HMGB1 overexpression, cellular responses to infection, and accumulation of viral particles. In our study, we chose a 24-hr infection period as it was sufficient for achieving HMGB1 overexpression without excessively compromising cell viability. Extending the infection beyond this point may have led to a more pronounced cytotoxic effect, making it important to carefully optimize infection conditions when using oncolytic viruses in research and potential clinical applications.
Our results from scratch assays showed no significant migration in HSV-HMGB1-infected CRC cells under both normoxic and hypoxic conditions, indicating that HMGB1 overexpression alone may not directly induce migration in CRC. It suggests that HMGB1 alone may not be the primary driver of migration in CRC cells, at least under the conditions tested in our study. However, this finding adds to the complexity of understanding the role of HMGB1 in cancer progression. It's important to note that cancer cell migration is a multifaceted process influenced by various factors, and our study focused on one specific aspect of this process.
The PPI network analysis revealed nine hub proteins, including UBC, NDRG1, ENO2, LGALS1, NDUFA4L2, ANGPTL4, WRNIP1, ALDOC, and PCK1. GO analysis demonstrated that these hub genes were significantly enriched in pathways related to gluconeogenesis, glycolytic process, and cellular response to hypoxia, among others. Additionally, our KEGG pathway analysis indicated significant enrichment in six pathways among the identified hub genes, shedding light on potential molecular mechanisms involved in the regulation of migration in CRC cells. Notably, among these enriched pathways, the PARP signaling pathway emerged as a promising candidate with implications in cancer cell migration. The involvement of PARP signaling in migration processes has been well-documented in various cancer types 31,32. The PARP signaling pathway presents an intriguing avenue for further research in the context of oncolytic virotherapy. Its involvement in DNA repair, immune regulation, and EMT makes it a potential target for inhibiting cancer cell migration 33.
Among the nine hub genes identified through PPI network analysis, we selected three genes including NDRG1, LGALS1, and ANGPTL4 that have been reported to play a role in cancer cell migration 34-36. The selection process was based on topological parameters, including degree and betweenness centrality, which provided valuable insights into the significance of these genes within the network. For further validation, we focused on three migration-related genes NDRG1, LGALS1, and ANGPTL4 and performed qRT-PCR analysis on HT29 cells infected with HSV-HMGB1 and HSV-ble under hypoxic and normoxic conditions. Consistent with the microarray dataset results, we observed differential expression of ANGPTL4, LGALS1, and NDRG1 under hypoxic conditions. ANGPTL4, LGALS1 and NDRG1 mRNA levels were upregulated in HT29 cells under hypoxic conditions. However, in HSV-HMGB1 and HSV-ble infected cells, the expression levels of these genes remained unchanged under both normoxic and hypoxic conditions. This suggests that HMGB1 overexpression does not significantly influence the expression of these migration-related genes.
ANGPTLs (angiopoietin-like proteins) are a family of proteins that are widely involved in metabolic disease and tumorigenesis 37. Intriguingly, the role of ANGPTL4 in cancer cell migration has been highlighted in previous studies 38-40. Notably, a study conducted by Zhang et al explored the transcription and protein expression patterns of ANGPTLs in CRC tissues in comparison to normal tissues 34. Their findings revealed a strong association between elevated ANGPTL expression and unfavorable oncological outcomes. Significantly, ANGPTL4 exhibited a significant positive correlation with CRC stage, implicating its role in promoting the proliferation and migration CRC cells 34.
NDRG1 plays pivotal roles in both CRC tumor growth and metastasis 35,41,42. Ma et al showed NDRG1’s involvement in the regulation of a Epithelial-Mesenchymal Transition (EMT), a pivotal process associated with cancer cell migration 35. When NDRG1 is suppressed, it triggers EMT in CRC cells. This activation of EMT is mediated through the NF-κB signaling pathway, a complex cellular mechanism involved in various processes including inflammation and cell migration 35. Galectin-1, the product of LGALS1, a homodimer comprised of 14 kD subunits, and belongs to a family of soluble galactoside-binding proteins 43,44. Research on LGALS1 highlights its pivotal role in the pathogenesis of CRC. This work has unveiled the significance of LYAR, a key regulator, in promoting CRC cell migration and invasion by upregulation of galectin-1 36.
In summary, our findings highlight the complex regulation of migration in CRC cells, which encompasses mechanisms dependent on hypoxia as well as those independent of HMGB1. The hub genes identified and the enriched pathways offer valuable targets for further exploration, contributing to a deeper understanding of the molecular pathways involved in CRC cell migration.
It is important to note that the microarray dataset we utilized was limited to HT29 cells under hypoxic and normoxic conditions and did not contain data regarding HSV-infected cells. Given the absence of suitable datasets for CRC cells under HSV infection conditions, we obtained and analyzed a microarray dataset from infected MRC5 cells. While we acknowledge that these cells differ from CRC cells, this dataset provided valuable insights into the pattern of migration-related genes after HSV infection, given the limitations of available data sources. Moreover, the direct translation of these findings to CRC cells and clinical practice requires further investigation using relevant CRC cell lines or patient-derived models. The data from MRC5 cells can serve as a starting point for understanding the broad principles of oncolytic virotherapy.
Utilizing the GSE124118 dataset, which was chosen due to its availability and compatibility, we sought to shed light on the impact of HSV-1 infection in a distinct cellular context. Upon examination of the three hub genes previously validated through real-time PCR, a consistent pattern emerged. In contrast to the response seen in hypoxia-conditioned cancer cells, where NDRG1, LGALS1, and ANGPTL4 exhibited distinct expression profiles, our real-time PCR analysis revealed that these genes remained remarkably stable in infected HT29 cells, regardless of whether they were under normoxic or hypoxic conditions. This intriguing observation underscores the unique interplay between HSV-HMGB1 and these genes in cancer cells, suggesting a potentially critical role in the context of virotherapy.
Conclusion :
In conclusion, our study not only sheds light on the intriguing interplay between HMGB1 and key migration-related genes in CRC cells, but also highlights the potential impact of oncolytic virotherapy in this context. Oncolytic virotherapy, such as HSV-HMGB1, could inhibits cancer cell migration through several mechanisms. These include direct targeting of cancer cells, immune system activation, modification of the tumor microenvironment and potential inhibition of migration-related signaling pathways. These speculative mechanisms suggest that oncolytic virotherapy could play a role in impeding cancer cell migration. While these ideas hold promise, it's important to emphasize that further research is required to validate these hypotheses and gain a comprehensive understanding of the involved mechanisms.
The stability of these genes NDRG1, LGALS1, and ANGPTL4 in HSV-HMGB1-infected cancer cells underlines the complex dynamics at play during virotherapy. The observed stability of NDRG1, LGALS1, and ANGPTL4 in HSV-HMGB1-infected cancer cells is likely the result of complex interplay between multiple regulatory mechanisms, signaling pathways, and the specific cellular context. Future research aimed at dissecting these mechanisms, possibly through transcriptomics, epigenomics, and functional studies, will provide valuable insights into the underlying biology and may uncover novel targets for therapeutic intervention.
While further investigations are warranted to fully elucidate these mechanisms, our research findings offer promise for the use of oncolytic virotherapy in CRC treatment, particularly in the context of limiting cancer cell migration. However, addressing the challenges associated with this approach, including optimizing immune responses, overcoming resistance, and ensuring affordability and accessibility, is crucial for realizing its full clinical potential. Continued research and clinical trials will be instrumental in advancing oncolytic virotherapy as a valuable tool in the fight against CRC and other malignancies. Moreover, the impact of hypoxia on cancer cell migration, as discussed in this study, has important clinical implications. It underscores the need to consider the tumor microenvironment, particularly hypoxia, when developing therapeutic strategies for CRC and other malignancies. Modulating the microenvironment and addressing hypoxia can be an integral part of future cancer treatment approaches.
In summary, this study delves into the intricate interplay between HMGB1, oncolytic virotherapy, and cancer cell migration, offering significant promise for cancer treatment. It points to several vital research directions: understanding the molecular mechanisms of HMGB1's effects, exploring HMGB1 as a therapeutic target, investigating synergies with other therapies, utilizing patient-derived models, validating findings in animal models, identifying relevant biomarkers, studying long-term treatment outcomes, assessing immune modulation, and working on strategies for translating preclinical findings into clinical practice. These research avenues hold the potential to enhance our comprehension of these mechanisms and, in the long run, lead to more effective therapies for colorectal cancer and other malignancies.
Ethical Statement :
Not applicable.
Conflict of Interest :
None declared.
Funding: This study was funded as Ph.D. student project by Pasteur Institute of Iran, Tehran.

Figure 1. Cell migration assays in Colorectal cancer cells under normoxic and hypoxic conditions. A) Scratch assays were performed on HT29, HCT116, and SW480 cells to assess migration under normoxic conditions. B) Scratch assays were performed on HT29, HCT116, and SW480 cells to assess migration under hypoxic conditions. Both panels (A) and (B), cells were either infected with HSV-HMGB1 to induce HMGB1 overexpression or with the control virus HSV-ble. The gap filling was examined at 24 hr post-scratch. No significant migration was observed in HSV-HMGB1-infected CRC cells under any tested conditions, including hypoxic environments. Similarly, no migration was detected in the control cells infected with HSV-ble under both normoxic and hypoxic conditions. Notably, migration was observed in uninfected CRC cells under hypoxic conditions, suggesting that hypoxia alone can induce migration in all three cell lines. The experiments were performed with six replicate, and representative images are shown. Error bars represent standard deviation. Image analysis was conducted using Image J software. *p<0.05, **p<0.01.
|
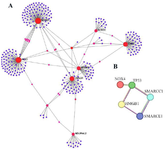
Figure 2. A) PPI network obtained from GSE9234 microarray dataset. Red, and purple circles represent genes. The area of each circle demonstrates the degree of the node in the network. The color of nodes is proportional to their BC values. B) PPI subnetwork with centrality of HMGB1.
|
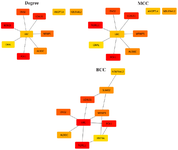
Figure 3. Results of algorithms from the Cytohubba. Top 10 Hub genes were screened by degree, MCC, and BC according to the Cytohubba plug-in. The more forward ranking is represented by a redder color.
|

Figure 4. DAVID functional GO analysis of (A) BP, CC, MF and (B) KEGG protein enrichment. DAVID, Database for Annotation, Visualization and Integrated Discovery; KEGG, Kyoto Encyclopedia of Genes and Genomes; GO, gene ontology; BP, biological processes; CC, cellular components; MF, molecular function. *p<0.05.
|

Figure 5. mRNA expression of LGALS1, ANGPTL4, and NDRG1 in HT29 cells infected with HSV-HMGB1 and HSV-ble under hypoxic and normoxic conditions were analyzed by reverse transcription-quantitative PCR. Gene expression levels were calculated based on Delta-Delta Ct relative quantification. Three biological replicates were performed. *p<0.05.
|
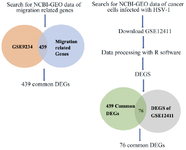
Figure 6. Overall diagram of data collection, processing, and analysis. Identification of 439 migration related DEGs from expression profile dataset (GSE9234). And Identification of 76 migration related DEGs from expression profile dataset (GSE12411).
|
![<p>Table 1. Summary of the nine selected hub proteins based on degree, MCC, and BC</p>
<p>Since there is no edge between the neighbors of the node, the MCC is equal to its degree. NDRG1: N-Myc Downstream Regulated 1; PCK1: Phosphoenolpyruvate Carboxykinase 1; LGALS1: Galectin 1; ENO2: Enolase 2; WRINP1: Werner Interacting Protein 1; ALDOC: <em>Aldolase, Fructose-Bisphosphate C; </em>NDUFA4L2: <em>NADH dehydrogenase [ubiquinone] 1 alpha subcomplex, 4-like 2; </em>UBC: Ubiquitin C; ANGPTL4: Angiopoietin-like 4.</p>](Images/Articles/60575/t1_small.png)
Table 1. Summary of the nine selected hub proteins based on degree, MCC, and BC
Since there is no edge between the neighbors of the node, the MCC is equal to its degree. NDRG1: N-Myc Downstream Regulated 1; PCK1: Phosphoenolpyruvate Carboxykinase 1; LGALS1: Galectin 1; ENO2: Enolase 2; WRINP1: Werner Interacting Protein 1; ALDOC: Aldolase, Fructose-Bisphosphate C; NDUFA4L2: NADH dehydrogenase [ubiquinone] 1 alpha subcomplex, 4-like 2; UBC: Ubiquitin C; ANGPTL4: Angiopoietin-like 4.
|
|